КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физические и физико-химические свойства биологических мембран
|
|
|
|
В живых клетках БМ пребывает в жидкокристаллическом состоянии. В жидком кристалле сочетаются свойства кристалла (дальний порядок организации и двулучепреломление) и жидкости (текучесть и образование капель), что приводит к возникновению качественно новых свойств. Термин «жидкий кристалл» существует в науке с конца XIX в. В1888 г. ботаник Ф. Райнитцер синтезировал эфир холестерина (холестерилбензоат) и изучил его свойства. Оказалось, что при температуре (Т) ниже 145 °С он был твердым кристаллом, при Т > 178 °С − прозрачной жидкостью, а в промежутке между этими температурами − мутной жидкостью. В следующем году физик О. Леман при исследовании мутной жидкости холестерилбензоата (при 145 < Т < 178 °С) в поляризационном микроскопе обнаружил анизотропию (типичное свойство кристалла) и назвал это промежуточное (между кристаллом и жидкостью) состояние холестерилбензоата жидкокристаллическим. Позднее оно было обнаружено у многих веществ при разных температурах и других параметрах среды (рН, росм и т. д.).
Бимолекулярный слой фосфолипидов, образующий биомембраны, в физиологических условиях (при температуре тела, нормальных рН и ионном составе интерстиция и цитозоля) представляет собой жидкий кристалл.
При понижении температуры и под влиянием ряда других факторов фосфолипидный каркас биомембран приобретает свойства твердого (димерного) кристалла, сохраняя бимолекулярную (в профиле) структуру. В обоих состояниях молекулы фосфолипида имеют гексагональную упаковку в плоскости биомембраны, но плотность их упаковки различна. Например, молекула лецитина занимает в твердокристаллическом состоянии поверхность в 0,46 − 0,48 нм2, а в жидкокристаллическом − 0,6 − 0,8 нм2. Чем больше двойных связей в неначищенных жирных кислотах, входящих в состав фосфолипидов, тем ниже температура, при которой они становятся твердыми кристаллами.
|
|
|
В физиологических условиях текучесть биологических мембран уменьшается при повышении в них содержания холестерина, ионов кальция и магния. Двухвалентные ионы в зависимости от концентрации нейтрализуют в той или иной степени отрицательный заряд на головках фосфолипидов и ослабляют их взаимное отталкивание, что приводит к более плотной упаковке молекул в биомембране. Местные анестетики (новокаин и родственные ему соединения) повышают степень текучести клеточных мембран, влияя на их жидкокристаллическое состояние. Оно изменяется при росте и развитии клеток, а также при некоторых патологических состояниях (раке, дистрофиях и др.).
Характерным свойством жидких кристаллов (ЖК) является их способность к фа зовым переходам, т. е. к преобразованию в твердые кристаллы (ТК) и возвращению в прежнее состояние (ЖК ←→ TK) при определенных условиях. В биологических мембранах фазовые переходы происходят в физиологических условиях под действием ряда агентов (раздражителей). Важно, что это может совершиться не во всем объеме мембраны, а в небольших ее участках (там, где появляются такие агенты).
Фазовый переход представляет собой кооперативный процесс. Он подчиняется закону «всё или ничего»: при плавном (градуальном) изменении силы раздражителя (S) физико-химические свойства биомембраны (вязкость, энергия активизации и другие) изменяются скачком. При S < Snop. сохраняется жидкокристаллическое состояние, а при S ≥ Snop небольшой участок биологической мембраны становится твердым кристаллом. Аналогично совершается обратный фазовый переход, когда твердая фаза сменяется жидкой (с более низкой вязкостью, в частности). Величина Snop для фазовых переходов в клеточных мембранах может быть очень малой. Они совершаются, например, при ничтожных концентрациях (менее 10-11 моль · л-1) некоторых физиологически активных веществ и фармакологических препаратов. Фактические данные о фазовых переходах в клеточных мембранах оживили давний интерес к гомеопатии.
|
|
|
Фазовый переход: ЖК ←→ TK в фосфолипидном каркасе существенно изменяет свойства ферментов, каналов, переносчиков и других функционально значимых компонентов биомембраны, находящихся в том ее участке, где совершается кооперативный процесс. Там изменяется проницаемость, нарушаются биохимические реакции, рецепторные и другие процессы, которые приводят к сдвигам в физиологическом состоянии организма. Кстати говоря, самим мембранным белкам также свойственно жидкокристаллическое состояние, и они могут испытывать фазовый переход.
Количество молекул, образующих участок биомембраны, где совершается кооперативный процесс, называется размером кооперативной единицы. В однородной среде он больше, чем в неоднородной. Белки и холестерин, встроенные в более или менее однородный фосфолипидный каркас, нарушают его однородность и тем самым уменьшают размер кооперативной единицы. Так обеспечивается высочайшая степень локальности фазовых переходов в клеточных мембранах.
Природа жидкокристаллического состояния биомембран обусловлена необычайно высокой подвижностью мембранных компонентов.
Подвижность молекулярных компонентов биомембран. Основными формами молекулярного движения в БМ являются латеральная миграция (перемещение молекул в плоскости мембраны, т. е. в пределах одной стороны бимолекулярного слоя) и вращение молекул вокруг собственной оси (например, родопсин вращается вокруг оси, перпендикулярной плоскости мембраны). Скорости обоих процессов примерно такие же, как скорость свободной диффузии в вязкой среде.
Большой свободой движения обладают липиды. Среднее время пребывания фосфолипидной молекулы в данном пункте мембраны не более 10-7 с. Следовательно, мембранные липиды за счет латеральной миграции беспрестанно меняются местами, причем каждая молекула меняет своих соседей миллион раз в секунду, передвигаясь со скоростью 5 − 10 мкм · с-1. В жидкокристаллической структуре молекулярные перемещения совершаются скачками. Между частотой (v) таких перескоков, площадью (S), занимаемой молекулой в БМ (например, у лецитина S = 0,6 − 0,8 нм2), и средним расстоянием (х−), проходимым молекулой за время t, установлены следующие соотношения: v=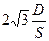 и х− = 4Dt, где D [м2 · с-1] − коэффициент латеральной миграции (диффузии) молекул. Его величина у мембранных фосфолипидов достигает (6 − 12) · 10-12 м2 · с-1.
и х− = 4Dt, где D [м2 · с-1] − коэффициент латеральной миграции (диффузии) молекул. Его величина у мембранных фосфолипидов достигает (6 − 12) · 10-12 м2 · с-1.
|
|
|
Примерно те же значения D характерны и для латеральной миграции некоторых мембранных белков. Однако у большинства из них частота перескоков меньше. Так, коэффициент латеральной диффузии антигенов по плазмолемме лимфоцитов не более 10-14 м2 · с-1, что соответствует скорости около 0,2 мкм · с-1. Изучение подвижности протеинов в БМ осложняется тем, что часть мембранных белков «заякорена» в определенных местах как гликопротеидами самих мембран, так и цитоплазматическими ультраструктурами (микротрубочками, микрофиламентами и т. п.). Есть и такие протеины, которые строго фиксированы в мембране (например, бактериородопсин в плазмолемме галобактерии).
Скорости вращательного движения мембранных молекул относительно нормали к поверхности мембраны довольно велики. Так, поворот на 1 радиан фосфолипид совершает примерно за 10-9 с, родопсин − за 10-6 с, цитохромоксидаза (фермент дыхательной цепи митохондрий) − за 10-4 с. Эта величина называется временем корреляции вращательного движения.
В отличие от вращения и латеральной миграции, перемещения молекул поперек мембраны (с одной стороны липидного бислоя на другую) совершаются очень редко. Такой вид молекулярного движения называют «флип-флоп» - перемещениями (или перескоками типа сальто-мортале). В искусственных мембранах «флип-флоп» - перемещения липидов происходят не чаще одного раза за несколько часов. Есть данные, что в БМ частота «флип-флопа» выше, но и там переходы молекул из слоя в слой совершаются гораздо реже, чем их латеральная миграция. Разница в вероятности этих видов молекулярного движения в клеточных мембранах обеспечивает постоянное поддержание их асимметрии, которая состоит в неодинаковом липидном и белковом составе их внутренней и наружной сторон.
|
|
|
В плазмолемме всех клеток млекопитающих наружная сторона насыщена холинфосфатидами (фосфатидилхолин, сфингомиелин), а внутренняя − аминофосфатидами (фосфатидилэтаноламин, фосфатидилсерин). По-видимому, вследствие асимметрии липидов в БМ ее наружная и внутренняя поверхности деформируются в неодинаковой степени при изменении температуры и под влиянием химических агентов. В результате таких воздействий может измениться кривизна клеточной мембраны. Еще более выражена асимметрия в распределении мембранных протеинов и углеводов. Внутренняя сторона БМ, как правило, свободна от углеводов.
Гликопротеиды сосредоточены преимущественно на наружной стороне плазмолеммы и обеспечивают рецепторную функцию. Напротив, основная часть ферментов, присущих плазматической мембране большинства клеток, встроена во внутреннюю половину липидного бислоя. Однако из этого правила есть исключения. Так, наружная сторона плазмолеммы кишечного эпителиоцита, обращенная в полость кишки, богата гидролитическими ферментами, которые обеспечивают важнейший элемент пищеварительного процесса − пристеночное пищеварение, открытое А.М. Уголевым в середине XX в. и называемое теперь мембранным пищеварением.
Асимметрия клеточных мембран имеет важное значение и в переносе через них различных веществ. Асимметричная ориентация белков и липидов, обеспечивающих мембранный транспорт, обусловливает векторные свойства биомембраны, т. е. однонаправленный перенос веществ через нее. Одни вещества (например, глюкоза) проходят только из полости кишки в кишечный эпителиоцит, но не идут в противоположном направлении. Продукты выделения транспортируются главным образом из клеток наружу.
Приведенные факты хорошо иллюстрируют основную особенность жидкокристаллического состояния − сочетание высокой упорядоченности с большой подвижностью молекулярных компонентов БМ. Диалектика взаимоотношений между этими, казалось бы, трудно сочетаемыми свойствами состоит в том, что сама лабильность жидких кристаллов обеспечивает поддержание стабильности образуемых ими структур в открытой системе при меняющихся условиях ее существования. Например, на разных стадиях развития клетки ферментный состав некоторых ее органоидов (в частности, эндоплазматической сети) должен изменяться. Эти изменения еще более выражены в неблагоприятных условиях существования клетки. Приспособление молекулярного состава БМ к таким условиям позволяет клетке сохранить жизнедеятельность. Говорят, что жидкокристаллическое состояние клеточных мембран способствует «затуханию беспорядка» в них вскоре после его возникновения. До сих пор не понято значение недавно обнаруженного феномена выравнивания липидного состава мембран разных типов в некоторых клетках злокачественных опухолей.
Лабильность БМ проявляется не только в значительной подвижности их молекулярных компонентов, но и в высоких темпах обновления, оцениваемых временем полужизни каждого из них. Среднее значение этого показателя различно у белков в разных мембранах: в плазмолемме и мембранах эндоплазматической сети − 50 ч, в митохондриальных мембранах − 110 ч, в ядерной мембране − 120 ч. Липиды также обновляются довольно быстро: у разных фосфолипидов время полужизни колеблется от 15 до 80 ч, а у холестерина − от 24 до 140 ч. Следовательно, в течение жизни клетки ее мембранные компоненты многократно обновляются. Однако это не приводит к замене целых мембранных систем, так как скорости обновления молекул разных типов в одних и тех же БМ варьируют в широких пределах.
Данные об изменчивости физико-химических свойств клеточных мембран позволяют считать их весьма динамичными структурами. Они могут перемещаться, исчезать, возникать вновь. В клетке непрерывно происходит своеобразный мембранный обмен.
Биологическим мембранам присущи и такие свойства, как значительная прочность на разрыв, упругость (эластичность), поверхностное натяжение, вязкость, электрострикция и флексоэлектрический эффект (способность генерации электрических потенциалов на мембране при ее деформации). Из перечисленных свойств два последних связаны с наличием поверхностного заряда на БМ.
Поверхностный заряд на клеточной мембране. Поверхностный заряд создается полярными головками фосфолипидов, гликопротеидами (главным образом карбоксильными группами сиаловой кислоты и аминокислотными остатками), гликолипидами. За счет этих веществ поверхность БМ заряжена отрицательно. Поверхностный заряд плазмолеммы играет важную роль в межклеточных взаимодействиях. Он способствует стабильности мембранных структур, а также связыванию ионов, находящихся в межклеточной среде. От поверхностного заряда плазмолеммы зависит ионный состав примембранных слоев межклеточной среды, что оказывает влияние на многие внутриклеточные обменные процессы.
Существование заряженных групп на БМ приводит к образованию диффузионного двойного электрического слоя, в котором фиксированный отрицательный заряд клеточной поверхности уравновешен положительным зарядом, создаваемым межклеточной средой за счет ионов. Разность потенциалов между частями двойного электрического слоя называют электрокинетическим потенциалом (или дзета-потенциалом). Это не что иное, как потенциал внешней поверхности БМ относительно интерстиция. Толщина двойного электрического слоя и величина дзета-потенциала зависят от природы электролита и концентрации его ионов. При уменьшении концентрации хлорида натрия в межклеточной среде в 200 раз толщина двойного слоя возрастает в 5 раз, а дзета-потенциал связан с этой толщиной экспоненциальной зависимостью. При высокой концентрации электролита дзета-потенциал стремится к нулю. Когда в межклеточной среде присутствуют двухвалентные катионы, избыток положительных зарядов может стать настолько значительным, что дзета-потенциал изменит свой знак. Снижение дзета-потенциала и, тем более, изменение его знака на противоположный сопровождается слипанием плазматических мембран соседних клеток. Так бывает при избытке Са2+ в межклеточной среде, а также при перестройках клеточных мембран, сопровождающихся сдвигами дзета-потенциала. Подобный эффект имеет некоторые особенности у клеток крови (например, эритроцитов), у которых дзета-потенциал может падать за счет нарушения не только солевого, но и белкового состава кровяной плазмы. Это явление лежит в основе изменения скорости оседания эритроцитов (СОЭ), измерения которой широко используют в диагностических целях. За счет дзета-потенциала имеет место электролиз клеток − в электрическом поле они движутся к аноду.
Дзета-потенциал, измеренный на поверхности у разных клеток, варьирует от -10 до -30 мВ. Его величина спадает в интерстиции и цитозоле по экспоненте с увеличением расстояния от наружной и внутренней поверхности БМ. Для оценки декремента (затухания) дзета-потенциала служит так называемый радиус экранировки Дебая − расстояние, на котором потенциал падает в е раз. В интерстиции он равен 0,8 нм, т. е. составляет примерно десятую часть толщины биомембраны. Следовательно, дзета-потенциал действует на очень малом расстоянии, но оказывает существенное влияние на размеры межклеточных пространств, противодействуя силам притяжения Ван-дер-Ваальса. Наименьшее расстояние между клетками составляет 10 − 20 нм. При таком расстоянии существует энергетическая яма во взаимодействии кулоновских и ван-дер-ваальсовых сил. Однако те ткани, клетки которых обладают большим дзета-потенциалом, имеют более протяженные (до 10 мкм) межклеточные промежутки. В большинстве тканей просвет между клеточными поверхностями составляет от 100 нм до 1 мкм.
В последние годы достигнуты немалые успехи в изучении поверхностного заряда как плазмолеммы, так и внутриклеточных мембран. Они связаны прежде всего с применением специальных люминофоров − флуоресцентных зондов, интенсивность свечения которых определяется величиной мембранного заряда.
Липидная часть БМ обладает свойствами диэлектрика. Диэлектрическая проницаемость гидрофобной зоны составляет 2,0 − 2,2, тогда как у гидрофильной зоны она гораздо выше (10 − 20). Электрическая емкость (С) полярных головок фосфолипидов достигает 30 мкФ · см-2, а жирнокислотные хвосты имеют С = 0,5 − 0,9 мкФ · см-2, причем она обратно пропорциональна числу атомов углерода в углеводородной цепи липида. Общая емкость клеточной мембраны не может превосходить наименьшую С ее компонентов, так как они образуют цепь с последовательным включением конденсаторов. Поэтому электрическая емкость БМ имеет порядок 1 мкФ · см-2.
Благодаря существованию в клеточной мембране заряженных групп, ей присуща выраженная электрострикция: по мере повышения трансмембранной разности потенциалов мембрана сжимается, что приводит к утончению гидрофобной зоны и, следовательно, к увеличению С клеточной мембраны. Возрастание дзета-потенциала сопровождается увеличением поверхности клетки.
Механические свойства биомембран. Механические свойства клеточных мембран весьма своеобразны. Величина модуля Юнга в поперечном направлении достигает 108 − 109 Па. Вместе с тем устойчивость к деформации сдвига примерно на 4 порядка меньше. Сила, достаточная для разрушения БМ на участке в 1 мкм, оценивается в 10-11 Н. Основной вклад в механическую прочность клеточной мембраны вносят белки, о чем свидетельствует сопоставление этих свойств у нее и искусственной липидной мембраны. Так, модуль упругости плазмолеммы эритроцита на 1 − 2 порядка больше, чем у искусственной липидной мембраны. Значительный вклад в повышение упругости и прочности эритроцитарной мембраны по сравнению с чисто липидной мембранной структурой вносят не только белки, встроенные в липидный бислой, но также довольно густая сеть, образованная мембранными белковыми молекулами и актиновыми нитями непосредственно под плазмолеммой в цитоплазме.
Вязкость биомембран. Биомембрана как жидкокристаллическая структура с присущей ей текучестью характеризуется определенной вязкостью, которая измерена методами ЭПР (электронный парамагнитный резонанс) и дифференциальной сканирующей микрокалориметрии (ДСК). Результаты измерений вязкости разными методами совпали. Вязкость БМ составляет от 0,03 до 0,1 Па · с (30 − 100 мПа · с = 30 − 100 сП), т. е. в 30-100 раз больше, чем у воды, и примерно такая же, как у оливкового масла.
При изменении температуры, молекулярного состава и других параметров БМ меняется ее вязкость вследствие возникновения фазового перехода: жидкий кристалл ←→ твердый кристалл (структура в виде относительно жестких вытянутых палочек). При температуре 310 − 311 К фосфолипиды с ненасыщенными углеводородными цепями пребывают в жидкокристаллическом состоянии, а при замене на полностью насыщенные цепи они образуют двухмерный кристалл. Регулируя молекулярный (прежде всего, фосфолипидный) состав клеточных мембран, живой организм может изменять их фазовые состояния при постоянной температуре и, напротив, сохранять жидкокристаллическое состояние при понижении температуры. Последнее свойственно бактериям и растениям, заменяющим в своих клеточных мембранах насыщенные липиды на ненасыщенные при охлаждении. Так они сохраняют жидкокристаллическое состояние, а значит, нормальную вязкость и зависящую от нее проницаемость мембран при низкой температуре окружающей среды.
Смена фосфолипидного состава БМ на «морозоустойчивый» присуща и пойкилотермным животным, которые используют этот механизм для адаптации к холоду. Строгое постоянство температуры тела гомойотермных животных избавляет их от необходимости осуществлять столь серьезные молекулярные перестройки своих клеточных мембран. Из сказанного следует, что одним из важных достоинств гомойотермии (постоянства температуры) является постоянное поддержание БМ в жидкокристаллическом состоянии, обеспечивающем стабильность транспорта веществ через них. Интересно, что у пингвинов температура нижних конечностей падает в дистальном направлении и, соответственно, мембраны клеток все более обогащаются ненасыщенными жирными кислотами (в тканях стопы их гораздо больше, чем в тканях бедра).
Вязкость БМ претерпевает существенные изменения при многих заболеваниях, а также под действием электромагнитных излучений (особенно ионизирующих), ряда фармакологических препаратов, гормонов и некоторых других факторов. Во многих случаях влияние на вязкость клеточных мембран опосредовано изменением содержания в них холестерина. При нормальной температуре тела повышение содержания холестерина в БМ увеличивает их вязкость, а понижение − уменьшает ее, В этом причина разжижения мембран в клетках злокачественных опухолей (например, лейкоцитарных мембран при лейкозе). Вязкость всех клеточных мембран падает при тиреотоксикозе, а также под действием наркотических веществ (например, хлороформа). Возможно, что наркотический эффект непосредственно связан с изменением физико-химических свойств биомембран. Так, головастик, помещенный в раствор хлороформа, утрачивает двигательную активность и способность реагировать на стимуляцию. Его удается «оживить», уплотнив клеточные мембраны путем повышения атмосферного давления. Однако при слишком высоком давлении головастик снова теряет подвижность и чувствительность. Их можно восстановить, добавив в среду наркотик. Известно, что дыхательная смесь, содержащая наряду с кислородом газ, оказывающий на организм наркотизирующее воздействие при атмосферном давлении, теряет этот эффект на большой глубине.
Приведенный пример позволяет сделать вывод, что жизнеспособность организма страдает как при понижении, так и при повышении мембранной проницаемости, которая, в свою очередь, зависит от вязкости БМ. Ее оптимальные значения поддерживаются в клеточных мембранах, пребывающих в жидкокристаллическом состоянии.
|
|
|
|
Дата добавления: 2014-01-05; Просмотров: 10293; Нарушение авторских прав?; Мы поможем в написании вашей работы!