КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Генетический анализ поведения с помощью спонтанных и индуцированных мутаций
|
|
|
|
Для изучения поведенческих признаков важно получить достаточно представительные выборки мутаций, влияющих на поведенческие признаки. При наличии таких выборок становится возможным выявить участие различных генов в контролировании данного признака, раскрыть характер их взаимодействия, подойти к механизмам генетического регулирования поведенческих особенностей.
Вопрос о необходимости получения в достаточных количествах нужных поведенческих мутаций у дрозофилы был поставлен С. Бензером в семидесятых годах. Бензер считал, что задача сводится к созданию аналогов селективных сред, где в качестве селектирующего фактора выступала бы сама поведенческая особенность мутантных мух. В ответ на соответствующий раздражитель они должны автоматически отделяться от остальной массы нормальных особей. Надежность выделения нужных мутантов повышают за счет многократного повторения процедуры сортировки особей по поведению.
Свои идеи Бензер реализовал при получении мутаций, индуцированных химическими мутагенами и затрагивающих фотореакцию. Был взят сильный алкилирующий агент этилметансульфонат, вызывающий в большом количестве точечные мутации типа замены пар оснований. Использовали методику «Muller-5», обычно применяемую для выделения рецессивных сцепленных с полом мутаций, а также некоторые другие методики и линии со сцепленными А-хромосомами у самок.
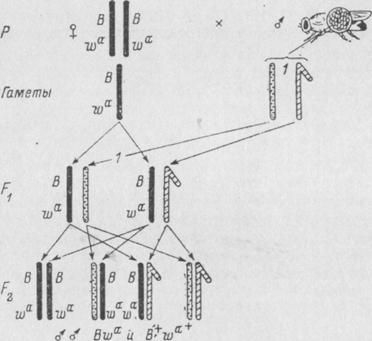
Рис. 1.38. Метод обнаружения рецессивных сцепленных с полом летальных
мутаций у дрозофилы (методика «Muller-5»)
В — ген полосковидных глаз; wa — ген абрикосового цвета глаз.
Эффективный способ выделения термочувствительных неврологических мутантов разработал в 1971 году Сузуки. Он, так же как и Бензер, использовал для индукции мутаций химические мутагены.
|
|
|
Высокоэффективную селективную среду для отбора физиологических мутантов с нулевой активностью алкогольдегидрогеназы (АДГ) разработали в 1972 году Софер и Хэткоф.
Таким образом, задача создания селективных методик для выделения физиологических и поведенческих мутаций у дрозофилы была решена. Но получение мутаций по данному признаку является лишь первым этапом генетической работы.
Второй этап изучения генетики поведения с помощью индуцированных мутаций заключается в установлении числа генов, к которым относятся выделенные мутации. Вероятны три ситуации:
1) все выделенные по данному признаку мутации будут результатом мутирования одного гена;
2) каждая отдельная мутация затронет свой локус,
3) мутирует несколько локусов, причем некоторые из них повторно.
Для определения принадлежности двух рецессивных мутаций к одному и тому же или разным генам используют функциональный тест на аллелизм. Для этого их объединяют в гетерозиготном состоянии в потомстве от скрещивания гомозиготных родителей:
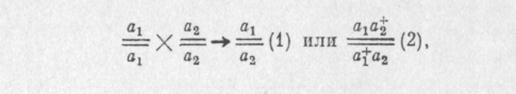
где % и at — мутации по данному поведенческому признаку.
Если у гетерозиготных особей F1 выявится аномальное поведение, то можно считать, что анализируемые мутации не комплементарны друг другу, т. е. являются аллелями одного гена, и их генотип следует записывать как (1). Если же у гетерозигот F1 обнаружится нормальное поведение, значит в анализ вовлечены комплементарные, взаимодополняющие мутации, которые относятся к разным локусам (2).
В работах Хота, Бензера, Сузуки было испытано на аллелизм по нескольку десятков рецессивных сцепленных с полом мутаций. Мутации, изменяющие фототаксис, возникли, как оказалось, в результате мутирования семи различных генов в Х-хромосоме, а температурозависимые мутации, вызывающие паралич, распределились по четырем генам.
|
|
|
Третий этап генетического анализа состоит в картировании локусов, по которым выделенные мутации распределены. Картирование новых мутантных генов, в том числе и влияющих на поведение, достигается с помощью тестерных линий, несущих в соответствующей хромосоме цепочку рецессивных и доминантных маркеров, т. е. легко опознаваемых и точно локализованных мутаций. В гибридном потомстве учитывают частоту рекомбинантных классов особей, несущих анализируемый признак и маркерные признаки. Необходимый результат может быть также достигнут с помощью тестерных линий, несущих серию делеций вдоль по длине хромосомы.
Гены, контролирующие данный поведенческий признак, могут распределиться на генетической карте самым различным образом. Например, Каплан с сотрудниками исследовал мутации, нарушающие локомоцию и четко проявляющиеся на провокационном фоне, — при эфирном наркозе. Тридцать полученных ими мутаций распределились по четырем локусам в Х-хромосоме на участке около 30% длины генетической карты, т. е. в 30 морганид. В другой работе 53 термочувствительные мутации, затронувшие 52 локуса, сосредоточились на одном участке, занимающем 13% длины генетической карты 3-й хромосомы.
Доведение генетического анализа до картирования отдельных генов, контролирующих физиологический или поведенческий признак, позволяет понять генетическую организацию этого признака.
Фенотипическую локализацию действия мутантного гена можно осуществить с помощью хромосомных мозаиков. В семидесятые годы Хотта и Бензером были получены мозаичные особи, несущие разные по величине нормальные и мутантные части тела.
В эксперименте использовались ранее полученные рецессивные мутации, распределенные по пяти локусам в Х-хромосоме и нарушающие фототаксис. Эти мутации путем кроссинговера порознь сочетались в одной хромосоме с рецессивными маркерами: у w sn f. Оппозиционная Х-хромосома несла нормальные аллели этих рецессивных мутаций, но включала структурную перестройку, придававшую ей форму кольца.
Такая необычная кольцевая хромосома уже на ранних эмбриональных стадиях развития имеет высокую вероятность претерпеть разрыв и в результате потеряться. Те ткани, в клетках которых сохранятся обе Х-хромосомы, будут по фенотипу нормальными. Те же ткани и органы, которые разовьются из клеток, утративших кольцевую Х-хромосому, окажутся мутантными, поскольку рецессивные мутации, будучи в гемизиготном состоянии, смогут теперь проявиться. Чем раньше в ходе онтогенеза произойдет элиминация кольцевой Х-хромосомы, тем больший участок тела будет иметь мутантный фенотип. При наличии маркеров, изменяющих окраску тела (у), глаз (w) и форму щетинок (sn и f), мутантные ткани легко различить на поверхности любого участка тела.
|
|
|
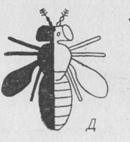
Рис. 1.39. Гинандроморф дрозофилы.
Левая сторона тела (черная) состоит из клеток с двумя Х-хромосомами, а правая (белая) - из клеток с одной хромосомой.

Рис. 1.40. Другие варианты строения гинандроморфов дрозофилы. Типы мозаичности фенотипа у гинандроморфных особей D.melanogaster по мутации hyperkinetic.
Известно, что в сложном глазу дрозофилы имеется около 800 омматидиев. (Омматидий — это структурная и функциональная единица фасеточного глаза насекомых. Омматидии сформировались в ходе эволюции из изолированных простых глазков при их интеграции в сложные, или фасеточные, глаза). Каждый омматидий включает 8 упорядоченно расположенных фоторецепторных клеток. Хотта и Бензер производили снятие электроретинограмм ЭРГ (это запись биоэлектрических потенциалов сетчатки глаза, возникающих при действии на нее света) у мозаичных особей, имеющих один глаз нормальный, а другой мутантный или же часть глаза нормальную, а другую мутантную. В отдельных случаях исследовали мозаиков с нарушением всего нескольких омматидиев. В результате была доказана высокая степень автономности фоторецепторных клеток глаза, функционирование которых определяется их собственными генотипическими особенностями.
Одновременно с работой Хотта и Бензера появилась работа Икеды и Каплана, которые применили метод получения мозаиков для нейрофизиологического исследования мутаций, вызывающих подергивание лапок у наркотизированных эфиром мух. Было изучено 77 гинандроморфов, несущих в Х-хромосоме соответствующую мутацию. С помощью микроэлектродной техники получили характеристики электрической активности отдельных нейронов торакального ганглия и доказали автономность отдельных мотонейронов в про-, мезо- и метаторакальных отделах торакального синганглия.
|
|
|
Без использования генетических подходов получение таких результатов вряд ли было бы возможным.
Исследование мозаичных особей позволило Хотта и Бензеру предложить способ вычленения так называемых фокусов — структур, ответственных за проявление поведенческих признаков.
Логика рассуждений была при этом следующая. В раннем эмбриогенезе дрозофилы, уже на стадии бластодермы, отдельные ее участки детерминированы. Из них формируются имагинальные диски, которые в свою очередь дают начало вполне определенным частям тела имаго. Если мозаицизм захватывает данный участок тела, что выявляется с помощью маркеров, и это сопровождается определенной поведенческой аномалией, то тем самым доказывается поражение нервной системы, мускулатуры или иных структур именно в данном участке. Таким путем было подтверждено, что мутации, нарушающие зрительную рецепцию и изменяющие электроретинограмму, затрагивают глаза или область, непосредственно к ним примыкающие. Мутация Hk 1 P нарушает функцию торакального ганглия. Сложный фокус мутации drop-dead (drd), вызывающей раннюю гибель мух, находится в церебральном ганглии. Билатеральные взаимосвязанные фокусы мутаций wings-up локализуются в районе торакса.
Получение мозаиков возможно лишь при утрате одной из Х-хромосом. Если же теряется одна из больших аутосом, то очень быстро наступает гибель особи. Холл и Кэнкель, сотрудники Бензера транслоцировали на Х-хромосому небольшие участки аутосом, нехватка по которым в гетерозиготном состоянии не приводит к летальному исходу. Эти исследователи в качестве маркеров использовали мутантные гены кислой фосфатазы и а-глицерофосфатдегидрогеназы, что дало возможность наблюдать мозаицизм тканей также и во внутренних органах.
При изучении генетики поведения используют и неврологические мутации. Например, Левин и Вимэн провели исследование морфологии летательных мышц и их иннервации, используя для этого мутацию stripe. Мутантные гомозиготные мухи не способны летать, их моторная активность редуцирована до кратковременных высокочастотных фибрилляций крыльев, однако видимых морфологических отклонений от нормы они не имеют. Основываясь на результатах микроэлектродной техники, авторы заключили, что ненормальное поведение мух stripe обусловлено дисбалансом в передаче импульсов, направляемых синергическим волокнам, и импульсов, направляемых мышцам-антагонистам.
Конопка и Бензер провели исследование циркадного ритма с помощью серии специально полученных мутаций. При применении химического мутагена этилметансульфоната (ЭМС) были получены три сцепленные с полом рецессивные мутации, изменяющие суточную цикличность вылупления имаго: per", per", per'. Все три мутации локализованы на коротком участке Х-хромосомы между генами zeste (1.0) и white (1.5).
Мутация null-period—arhythmic (per") обусловливает сравнительно равномерное распределение частот от рождения мух в течение суток. Мутация short-period (per') контролирует вполне четкий ритм, но не при 24-часовом, а при 19-часовом цикле. Мутация long-period (per1) удлиняет цикл вылупления до 28 часов.
В лаборатории Бензера в 1976 году была получена сцепленная с полом мутация dunce («тупица»). Особенность этих мутантов заключается в неспособности к выработке условных рефлексов избегания на специфический запах при подкреплении электрическим током. В то же время их обонятельные реакции, так же как другие физиологические и поведенческие особенности, сохраняются нормальными.
Получены мутации, обусловливающие резистентность к таким нейротропным и психотропным веществам, как амфетамин, барбитал, бидрин, физостигмин, кофеин, теофиллин и пикротоксин. Эти мутации представляют для исследования метаболизма нейромедиаторов.
Получение мутаций, влияющих на физиологические и поведенческие признаки, открывает благоприятные возможности для их последующего углубленного исследования. Генетический анализ таких мутаций может быть осуществлен в полной мере, вплоть до картирования на генетической карте. С их помощью может быть раскрыта система генетического контроля физиологических признаков.
Вклад мутационного анализа поведенческих признаков в генетику поведения является весьма значительным.
Генетический анализ поведения с помощью исследования сравнительно–генетических и селекционных методов.
Селекционный и сравнительно-генетический методы являются традиционными для генетики поведения, поскольку именно на них основывались первые представления о наследственной обусловленности поведенческих признаков.
Сравнительно-генетический метод заключается в выявлении различий среди особей, выращенных в идентичных условиях, но принадлежащих разным генетическим совокупностям, репродуктивно изолированным, например видам, популяциям или линиям.
Селекционный метод заключается в отборе для размножения в ряду поколений особей с определенными чертами поведения. Двусторонний отбор на альтернативные варианты признака (например, отрицательный или положительный фототаксис) или на разную степень проявления признака (высокую или низкую активность) позволяет не только практически вычленить из общей фенотипической изменчивости признака долю наследственно обусловленной вариабельности, но и получить линии, четко различающиеся по интересующему признаку.
В результате селекционного и сравнительно-генетического методов подбирают пары линий, контрастные по поведению, для их дальнейшего изучения гибридологическими, физиологическими и биохимическими методами.
Инбредные и селектированные линии имеют значительно более сложные различия, чем мутантные, выделенные из материнской линии в результате индуцированного мутагенеза, поэтому гибридологический анализ инбредных и селектированных линий, как правило, не доводят до локализации генов, ответственных за линейные различия.
Однако ограничить исследования в генетике поведения использованием только мутационных моделей не представляется возможным. Для решения ряда важных проблем в этой области селекционные модели более адекватны, нежели мутационные, например, при выяснении степени генетической обусловленности индивидуальной изменчивости поведения и адаптивного значения отдельных поведенческих особенностей, изучения популяционного полиморфизма поведенческих признаков, выявления роли поведения в микроэволюционных процессах.
Из двух традиционных подходов — сравнительно-генетического и селекционного — последний, более перспективен для модельных экспериментов по генетике поведения дрозофилы, поскольку позволяет вести исследования по интересующим свойствам поведения и получать линии с желаемой степенью контрастности.
Особое место в сравнительно-генетических исследованиях занимают сравнительно-видовые сопоставления. Они не претендуют на роль модельных, так как ограничивают применение гибридологического анализа: у дрозофилы, как у других организмов, межвидовые скрещивания редко дают жизнеспособное и фертильное потомство. Такого рода опыты преследуют специальные цели: оценку генетического расстояния между видами, изучение механизмов их изоляции, выяснение видовых особенностей образа жизни, выявление специфических и перекрывающихся экологических потребностей совместно обитающих видов.
Из всех признаков поведения в сравнительно-видовых исследованиях наибольшее внимание было уделено половому поведению. Особенности полового поведения были изучены у 101 вида рода Drosophila, относящимся к различным систематическим группам, что внесло определенный вклад в эволюционную систематику данного рода. Для большинства видов дрозофилы установлена более активная роль самок в избирательности спаривания, однако, в недавнее время обнаружен вид, где ведущую роль в этом отношении играет самец
Виды дрозофилы различаются по степени зависимости полового поведения от освещения, а также по способности самцов ухаживать за безголовыми самками.
В некоторых случаях благодаря скрещиваемости близкородственных видов группы D. paulistorum, а также D. simulans и D. melanogaster, D. pseudoobscura и D. persimilis, D. auraria и D. triauraria удалось осуществить элементарный генетический анализ видовых особенностей поведения с получением гибридов первого поколения.
Методом селекции можно повысить эффективность межвидовых скрещиваний, снизить их физиологическую изоляцию. Однако, при использовании такого приема для повышения разрешающей способности генетического анализа видовых признаков надо удостовериться, что селекция не затронула анализируемые признаки.
В опытах по генетике поведения используют такие стандартные линии как «Орегон», «Кантон-С», а также «Флорида», «Эдинбург» и несколько других, более редких линий. Дополнительным источником уникальных инбредных генотипов для сравнительно-генетических исследований у дрозофилы могут служить обширные наборы мутантных линий.
Была успешной селекция мышей и крыс на степень выраженности ряда поведенческих признаков. Осуществлены селекционные программы, которые охватили практически весь диапазон признаков поведения, показавших генетическую изменчивость, — от скорости проведения нервного возбуждения в нервно-мышечном синапсе до высокой и низкой способности к обучению. В то же время с помощью селекции формируются линии, которые состоят из животных, сходных по выраженности исследуемого признака и иногда ряда других признаков, но не обладающих генетической однородностью. Это затрудняет использование их в целях идентификации генов, влияющих на данный признак.
В ряде случаев удалось провести работу по выведению инбредных селектированных линий, т.е. полностью гомозиготных животных с контрастными в случае двух линий величинами интересующего исследователей признака. Инбридинг начинают после того, как получены неперекрывающиеся межлинейные различия по интересующему признаку. Таких линий немного. В качестве примера можно привести инбредные линии мышей Short- и Long sleep, у которых после гипногенной дозы этанола развивается либо короткий, либо длинный период сна. Основную же массу составляют инбредные линии мышей и крыс, выведенные ранее, до начала активных исследований в области генетики поведения. Такие линии были необходимы для исследований по иммунологии, гистосовместимости, онкологии.
Основными отличиями инбредных линий от генетически гетерогенных аутбредных, является изогенность, гомозиготность, фенотипическое подобие, долговременная стабильность, возможность идентификации.
Поскольку вероятность резких различий по поведению у случайно взятых линий невелика, обычно одновременно обследуют несколько линий. Если к поведенческим оценкам инбредных линий добавить оценки их гибридов, то появляется возможность разложить генетическую компоненту изменчивости признака на ее составляющие: изменчивость, зависящую от аддитивного действия генов, а также зависящую от аллельного и неаллельного взаимодействия генов, т. е. соответственно от доминирования и эпистаза.
Для оценки этих параметров разработан специальный метод диаллельного анализа. Метод основывается на представлениях о полигенном наследовании поведенческих признаков и на допущении наличия только двух аллелей для каждого локуса. Исходным материалом для математического анализа служат данные диаллельного скрещивания, т. е. всех возможных комбинаций между выбранными линиями. Диаллельный анализ, позволяет оценить нижний предел количества генов, влияющих на признак, определить их аддитивный эффект, степень и направление доминирования, охарактеризовать родительские линии по общей и специальной комбинативной способности и по относительному содержанию в них аллелей с рецессивным или доминантным действием, а также выявить различия в реципрокных скрещиваниях.
Диаллельный анализ является прекрасным методом для формально-генетического изучения изменчивости по признакам поведения в популяциях, для выяснения генетической организации признака на уровне популяции. Однако для анализа генетической детерминации признака на организменном уровне у дрозофилы существуют более тонкие и адекватные методы, а именно выделение и накопление мутантных генов, влияющих на признак, в селектируемых линиях с последующей их локализацией в хромосомах.
Глава 2. Пути реализации генетической информации на уровне поведения
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 765; Нарушение авторских прав?; Мы поможем в написании вашей работы!