КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Эндоплазматическая сеть
|
|
|
|
Эндоплозматическоя сеть (ЭПС) – органелла, обеспечивающая синтез углеводов, липидов и белков, а также начальные посттрансляционные изменения последних. Она имеет мембранное строение и состоит из системы уплощенных, удлиненных, трубчатых и везикулярных образований. Название органеллы обусловлено характером связи этих элементов друг с другом, образующих в цитоплазме непрерывную трехмерную сеть, элементы которой лишь на отдельных срезах могут иметь вид изолированных структур. Мембрана ЭПС тоньше, чем плазмолемма и содержит более высокую концентрацию белка, что связано с наличием в ней многочисленных ферментных систем. Степень развития ЭПС и особенности ее строения варьируют в различных клетках и зависят от их функции. Выделяют две разновидности ЭПС: гранулярную ЭПС (грЭПС) и гладкую, или агранулярную ЭПС (аЭПС), которые связаны (друг с другом в области перехода, называемой переходной (транзиторной ЭПС (рис.3-7)
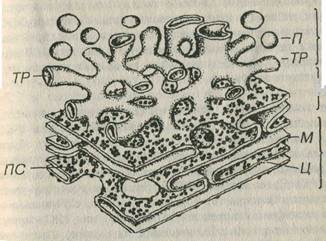
Рис 3-7. Эндоплаэматическая сеть. грЭПС: ПС - полисомы, М - мембрана, Ц- цистерны; аЭПС: ТР - трубочка, П - пузырьки; пЭПС - переходная ЭПС.
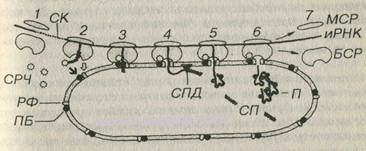
Рис 3-8 Синтез белка на гранулярной эндоплазматической сети. БСР -большая субъединица рибосомы, МСР - малая субъединица рибосомы, РФ - рибофорины СРЧ - сигнал-распознающая частица, ПБ - причальный белок, СК - сигнальные кодоны (иРНК), СП - сигнальный пептид, СПД - сигнальная пептидаза, П - пептид (продукт синтеза). Светлая стрелка - связывание БСР с РФ, темная стрелка - связывание СРЧ с ПБ.
Гранулярная ЭПС обеспечивает (1) биосинтез всех мембранных белков и белков, предназначенных для экспорта из клетки, и (2) начальное гликозилирование и посттрансляционные изменения белков молекул. Гранулярная ЭПС образована уплощенными мембранными цистернами и трубочками, на наружной поверхности которых располагаются рибосомы и полисомы, придающие мембранам зернистый (гранулярный) вид (см. рис. 3-7 и 3-8), что и отражено в названии органеллы. Мембраны грЭПС содержат особые белки, которые обеспечивают (1) связывание рибосом и (2) уплощеше цистерн. Полость грЭПС со держит рыхлый материал умеренной плотности (продукты синтеза) и сообщается с перинуклеарным пространством (см. ниже). Благодаря грЭПС происходит отделение (сегрегация) вновь синтезированных белковых молекул от гиалоплазмы.
|
|
|
Синтез белка на грЭПС начинается на свободных полисомах, которые в дальнейшем связываются с мембранами ЭПС (см. рис. 3-8). Н первом этапе взаимодействия иРНК с рибосомами происходит образование особого сигнального пептида (длиной 20-25 аминокислот), связывающегося с рибонуклеопротеидным комплексом – сигнал-распознающею частицей (СРЧ). Присоединение СРЧ к сигнальному пептиду угнетает дальнейший синтез белка до тех пор, пока комплекс СРЧ-полисома не свяжется со специфическим рецептором на мембране ЭПС - причальным белком (docking protein в англоязычной литературе). После связывания с рецептором СРЧ отделяется от полисом, что разблокирует синтез белковой молекулы. В мембране грЭПС имеются интегральные рецепторные белки рибофорины, обеспечивающие прикрепление больших субъединиц рибосом. Эти белки не диффундируют в область аЭПС и формируют гидрофобные каналы в мембране, служащие для проникновения вновь синтезированной белковой цепочки в просвет грЭПС что, наряду с рибофоринами, способствует удержанию рибосом на поверхности мембран грЭПС.
В просвете грЭПС сигнальный пептид отщепляется особым ферментом сигнальной пептидазой, которая располагается на внутренней поверхности мембраны. В ходе продолжающейся трансляции внутри цистерны грЭПС накапливается белок, который приобретает вторичную и третичную структуру, а также подвергается начальным посттрансляционным зменениям – гидроксилированию, сульфатированию и фосфорилированию. Наиболее важным из этих изменений является гликозилирование – присоединение к белкам олигосахаридов с образованием гликопротеинов, которое происходит перед секрецией или транспортом большинства белков к другим участкам внутри клетки (комплексу Гольджи, лизосомам или плазмолемме). В отличие от них, растворимые белки гиалоплазмы не гликозилированы. Гликозилирование обеспечивается связанным с мембраной ферментом гликозилтрансферазой, переносящим олигосахарид.
|
|
|
Хотя грЭПС присутствует во всех клетках (за исключением спермиев), степень ее развития существенно варьирует. Она особенно хорошо развита в клетках, специализирующихся на белковом синтезе, например, в эпителиальных железистых клетках ацинусов поджелудочной железы (вырабатывающих пищеварительные ферменты), фибробластах (синтезирующих коллаген и ряд других белков), плазматических клетках (продуцирующих иммуноглобулины). Для всех этих клеток характерна выраженная базофилия цитоплазмы в области расположения элементов грЭПС. В нейронах отдельным компактным скоплениям цистерн грЭПС на светооптаческом уровне соответствуют очерченные участки базофилии цитоплазмы, которые в совокупности называются хроматофильной субстанцией или тельцами Ниссля.
Агранулярная (гладкая) ЭПС представляет собой трехмерную замкнутую сеть мембранных анастомозирующих трубочек, канальцев, цистерн и пузырьков диаметром 20-100 нм, на поверхности которых рибосомы отсутствуют (см. рис. 3-7), что определило ее название. Соответственно, на мембранах аЭПС отсутствуют рецепторы, связывающие субъединицы рибосом (рибофорины). Предполагают, что аЭПС образуется в результате формирования выростов грЭПС, мембрана которых утрачивает рибосомы.
Функции аЭПС включают: (1) синтез липидов, в том числе мембранных (ферменты липидного синтеза располагаются на наружной – обращенной в сторону гиалоплазмы - поверхности мембраны аЭПС), (2) синтез гликогена, (3) синтез холестерина, (4) детоксикацию эндогенных и экзогенных веществ, (5) накопление ионов Са2+, (6) восстановление кариолеммы в телофазе митоза (эта функция оспаривается авторами, считающими, что кариолемма восстанавливается за счет мембранных пузырьков, на которые она ранее распалась). Помимо указанных основных функций, в некоторых типах клеток аЭПС выполняет ряд дополнительных – например, в мегакариоцитах (гигантских клетках костного мозга) ее элементы образуют демаркационные каналы, разделяющих формирующиеся тромбоциты.
|
|
|
Способность аЭПС к накоплению ионов Са2+ обусловлена наличием (1) кальциевого насоса в ее мембране, который обеспечивает транспорт этих ионов из гиалоплазмы внутрь цистерн аЭПС; (2) кальций-связывающих белков (кальсеквестрина в мышечных клетках, кальретикулина - преимущественно в немышечных и др.), которые в просвете цистерн образуют комплекс с ионами Са2+ и (3) кальциевых каналов в мембране аЭПС, которые осуществляют выведение Са2+ в гиалоплазму. Механизмы действия кальциевых каналов неодинаковы в клетка разных типов. Функция накопления ионов Са2+ особенно выражена в мышечных клетках, в которых специализированная аЭПС (именуема саркоплазматической сетью) обеспечивает мышечное сокращение путем накопления и выделения значительных количеств ионов Са2+, связывающихся с особыми белками.
Обычно аЭПС в вдтоплазме занимает меньший объем, чем грЭПС однако она очень хорошо развита в клетках, синтезирующих стероиды триглицериды и холестерин. Так, аЭПС занимает значительную часть объема цитоплазмы в клетеах, которые активно продуцируют стероидные гормоны (клетки коркового вещества надлочечника, интерстици| альные гландулоциты яичка (клетки Лейдига), клетки желтого тела яичника (лютеоциты) и др. Она также хорошо развита в клетках печенй (гепатоцитах), где ее ферменты участвуют в процессах окисления, конъюгации и метилирования, которые обеспечивают нейтрализацию и детоксикацию ряда гормонов и вредных вещеста (алкоголя, инсектицидов и др.).
Переходная (транзитортя) ЭПС - участок перехода грЭПС в аЭПС у формирующейся поверхности комплекса Гольджи. В области переходной ЭПС трубочки распадаются на отдельные фрагменты, образующие окаймленные транспортные пузырьки, которые переносят материал из ЭПС в комплекс Гольджи (рис. 3-9),
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 4397; Нарушение авторских прав?; Мы поможем в написании вашей работы!