КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Вплив зовнішніх факторів на фотосинтез. Показники фотосинтезу
Фотосинтез характеризується швидкістю або інтенсивністю. Інтенсивність фотосинтезу вимірюється кількістю міліграмів С02, який поглинений одним квадратним дециметром листової поверхні за 1 годину. Для її вимірювання визначають зменшення кількості вуглекислого газу в замкнутій камері, в якій розміщено листок. Одночасно разом з визначенням інтенсивності фотосинтезу визначають в темноті інтенсивність дихання цих клітин. Кількість С02, що вивільняється при диханні, додають до його кількості, що поглинена при освітленні. При цьому вважають, що інтенсивність дихання клітин на світлі і в темноті однакова. Однак, ці по¬правки не можуть точно оцінити дійсний фотосинтез, тому що при затінені листка виключається не тільки фотосин¬тез, але й гліколатний цикл, а також тому, що дихання в дійсності трохи залежить від світла.
Інтенсивність фотосинтезу можна виміряти і по кількості кисню, що виділяється с квадратного дециметра листової поверх¬ні за 1 годину. Величини інтенсивності фотосинтезу, що розраховані по кількості поглиненого вуглекислого газу і по кількості виділеного кисню, можуть не співпадати, тому що С02 приймає участь в темновій, а кисень утворюється в світловій фазі фотосинтезу. Співвідношення об’єму виділеного кисню до об’єму поглиненого вуглекислого газу називається фотосинтетичним коефіцієнтом. При утворенні вуглеводів, фотосинтетичний коефіцієнт дорівнює одиниці (602: 6С02 = 1). При утворенні білків він дорівнює 1,25, а при утворенні ліпідів – 1,44. Середня величина фотосинтетичного коефіцієнта у різних рослин дорівнює 1,04.
Величина фотосинтетичного коефіцієнта залежить від умов освітлення і мінерального живлення. Освітлення рослин си¬нім світлом і гарне постачання азоту збільшує синтез білків, таким чином фотосинтетичний коефіцієнт збільшується.
Для характеристики фотосинтезу використовують такі показники: квантова витрата, квантовий вихід і асиміляційне число. Квантова витрата фотосинтезу – це співвідношення числа по-глинених квантів світла до числа асимільованих молекул дво¬окису вуглецю. Обернена величина отримала назву квантового виходу. Асиміляційне число – це співвідношення кількості поглиненого вуглекислого газу до кількості хлорофілу, який міститься в листку.
Кількість грамів сухої речовини, яка накопичена в рослині за добу в перерахунку на 1 м2 листової поверхні, характеризує чисту продуктивність фотосинтезу (ЧПФ).
Вплив світла. Прямий вплив світла на фотосинтез, його субстратна роль обумовлена тим, що воно є джерелом енергії. Опосередковано світло впливає завдяки регуляції ширини продихових щілин, регулюючи дифузію СО2 в лист. Таким чином регулює як темнову так і світлову фази.
Залежність інтенсивності фотосинтезу від освітленості виражається логарифмічною кривою – світловою кривою фотосинтезу. По осі абсцис відкладають освітленість, а по оси ординат – інтенсивність фотосинтезу, яку вимірюють по мірі збільшення інтенсивності світла на одну і ту ж величину.
При збільшенні інтенсивності світла від повної темноти до 0,2-0,3 повного сонячного освітлення швидкість фотосинтезу зростає прямолінійно, потім уповільнюється і, при підвищенні освітленості більше 0,5-0,6 від повного сонячного освітлення - не викликає змін інтенсивності фотосинтезу. Такий стан називають станом світлового насичення. У рослин помірної зони світлове насичення наступає при освітленості 20-40 тис. лк.
При слабкій освітленості дихання відбувається швидше за фотосинтез, прилади реєструють виділення СО2, тому початок світлової кривої розташований нижче осі абсцис. Згодом швидкість фотосинтезу стає рівною швидкості дихання. При цьому вуглекислий газ ні виділяється, ні поглинається. Освітленість, при якій інтенсивність фотосинтезу дорівнює інтенсивності дихання, називають світловою компенсаційною точкою (світлова крива перетинає вісь абсцис).
При прямій пропорційності між інтенсивністю фотосинтезу і збільшенням інтенсивності світла, швидкість фотосинтезу визначається швидкістю світлової фази. В стані світлового насичення швидкість фотосинтезу лімітують темнові процеси і надходження СО2.
Подальше збільшення освітленості може призвести до зниження інтенсивності фотосинтезу, а в умовах надлишку освітлення порушується синтез пігментів, що інігібують хімічні реакції. В природних умовах недостатня освітленість листків часто обмежує фотосинтез.
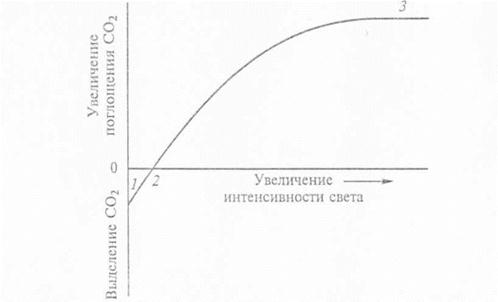
Залежність інтенсивності від освітленості: 1-швидкість виділення СО2 в темноті (швидкість дихання); 2-комненсаційна точка фотосинтезу; 3-стан світлового насичення.
Вплив вуглекислого газу. Вуглекислий газ є джерелом вуглецю для синтезу органічних речовин (субстратна роль) та регулює ширину продихових щілин.
В лабораторних умовах залежність інтенсивності фотосинтезу від концентрації С02 виражається логарифмічною кривою, подібною до світлової кривої фотосинтезу і називається вуглекислотною кривою фотосинтезу. При підвищенні концентрації С02 інтенсивність фотосинтезу спочатку зростає швидко, а потім повільніше і, згодом, збільшення кількості вуглекислого газу не викликає посилення фотосинтезу. Такий стан рослин називають вуглекислотним насиченням. Він наступає при концентраціях С02 в повітрі 0,1-0,3 %. При концентрації С02 0,005 % у С3-рослин і 0,0005 % у С4-рослин, швидкість фотосинтезу дорівнює швидкості дихання. Концентрацію С02, при якій наступає рівняння цих процесів, називають вуглекислотною компенсаційною точкою.
Швидкість асиміляції С02 в процесі фотосинтезу залежить від швидкості його надходження в хлоропласти з атмосфери, що визначається швидкістю дифузії крізь продихи в клітини.
Двоокис вуглецю поглинається з ґрунтового повітря коренями (темнова фіксація С02). Тут відбувається карбоксилювання органічних кислот (ФЕП). Такі кислоти транспортуються в листки, там декарбоксилюються, С02 використовується для фотосинтезу.
Надходження в рослини вуглекислого газу постійно лімітує фотосинтез. При вирощуванні С3-рослин в теплицях в атмосфері, збагаченій вуглекислим газом, врожай збільшується в 2-5 раз, тому що великі концентрації С02 пригнічують гліколатний цикл фотосинтезу.
Однак, вирощувати рослини в атмосфері, збагаченій С02, можна тільки на сильно освітлених місцях.
Вплив кисню. У більшості рослин зниження концентрації кисню с 21 % (звичайна в атмосферному повітрі) до 1-3 % активує фотосинтез. Це пояснюється тим, що фотосинтез виник в умовах низького вмісту кисню в атмосфері. 90-95 % кисню атмосфери Землі фо-тосинтетичного походження.
Пригнічення фотосинтезу киснем називають ефектом Варбурга. Ефект Варбурга виявлений у всіх С3-рослин; у С4-рослин при звичайній концентрації кисню відсутній. Цей ефект пояснюється: по-перше, при великій концентрації кисню починає проходити гліколатний цикл. При знижені концентрації кисню до 2-3 % гліколат не утворюється, і ефект Варбурга зникає; по-друге, при великій концентрації кисню може відбуватись псевдоциклічне фотофосфорилювання: електрони в світловій фазі транспортуються не до НАДФ+, а до кисню повітря. В результаті кисень відновлюється, утворюється перекис водню, який розкладається на воду і атомарний кисень. Кількість відновленого кисню дорівнює його кількості, що утворюється при фотолізі води. В атмосферу він не вивільняється. Зменшення кількості НАДФ·Н2 пригнічує темнову фазу фотосинтезу. В цьому процесі приймають участь обидві фотосистеми. Низький вміст або відсутність кисню або збільшення його концентрації до 25-30 % пригнічують фотосинтез.
Температура впливає в основному на темнову фазу фотосинтезу, регулюючи активність ферментів і швидкість дифузії вуглекислого газу. При підвищенні температури вище оптимального значення порушується фотосинтетичне фосфорилювання. Температура впливає на швидкість фотосинтезу і опосередковано, змінюючи швидкість відтоку асимілятів з листової пла¬стинки в інші органи, а накопичення асимілятів в листовій пластинці уповільнює фотосинтез.
Залежність інтенсивності фотосинтезу від температури зображують у вигляді одновершинної температурної кривої, на якій виділяються три кардинальні точки: мінімальна, оптимальна і максимальна температури.
У тропічних і субтропічних рослин фотосинтез починається при температурі +5...+7 °С, а у більшості рослин помірної зони — біля 0°С і тільки у деяких хвойних (сосна, ялина) — при від‘ємних температурах: -2...-3°С і навіть -15°С.
Поглинання і відновлення двоокису вуглецю у всіх рослин з підвищенням температури спочатку збільшується. У більшості рослин помірної зони найбільша інтенсивність фотосинтезу спостерігається при температурі 20-28°С, а у С4-рослин – при 35-45 і навіть при 50°С. При подальшому підвищенні температури швидкість фотосинтезу зменшується, але зростає інтенсивність дихання. Температуру, при якій інтенсивність фотосинтезу дорівнює інтенсивності дихання, називають температурною компенсаційною точкою. При такій температурі в рослині не відбувається накопичення сухої речовини. На температурну компенсаційну точку впливають освітленість і вміст вуглекислого газу в атмосфері. Для С3-рослин помірної зони максимальними є температури 40-50°С, а для тропічних видів 50-60°С. Під дією високих температур порушується структура хлоропластів. При мінімальних температурах повільно проходить темнова фаза фотосинтезу, при максимальних – інактивуються ферменти.
Положення кардинальних точок на температурній кривій залежить від тривалості дії даної температури.
При підвищенні температури на 10°С в межах від 0 до 25-35°С в залежності від виду рослини інтенсивність фотосинтезу збільшується в 2-3 рази (правило Вант-Гоффа). При подальшому підвищенні температури вона знижується.
Вплив оводненості тканин. Вода приймає участь в обох фазах фотосинтезу, виконуючи субстратну і регуляторну роль. Субстратна роль води полягає в тому, що під час светлової фази вона є донором водню для відновлення НАДФ. Для цього використовується приблизно 1 % поглиненої води.
Вода є джерелом водню для відновлення двоокису вуглецю, середовищем для всіх хімічних реакцій, активує ферменти. Завдяки випаровуванню відбувається регуляція температури тканин, від якої залежить швидкість темнової фази фотосинтезу. В умовах оптимальної кількості води синтез речовин відбувається швидше, ніж розпад. Кількість води впливає на швидкість відкладання первинного крохмалю і навіть на структуру і розташування тилакоїдів в стромі хлоропласта. Від кількості води в замикаючих клітинах залежить ступінь відкритості продихів, а від тургорного стану листків – їхнього розташування по відношенню до сонячних променів.
Залежність інтенсивності фотосинтезу від кількості води в тканинах виражається на графіку одновершинною кривою, яка має три кардинальні точки – мінімуму, оптимуму і максимуму. 85-87% води в клітинах вважається оптимальною величиною для фотосинтезу. Тому, максимальний фотосинтез у більшості наземних рослин відбувається при невеликому водному дефіциті. При втраті 50% води фотосинтез повністю зупиняється внаслідок закривання продихів. Одночасно при сильній дегідратації клітин порушується структура хлоропластів. Причинами зниження інтенсивності фотосинтезу також є зміни активності ферментів, дегідратація кутикули, стінок епідерми і мембран, в результаті зменшується проникність клітин для вуглеки¬слого газу. При нестачі води погано відбувається циклічне і нециклічне фосфорилювання.
При високій оводненості клітин (більше 87%), при відкритих продихах, інтенсивність фотосинтезу також знижується, завдяки порушенню дифузії С02, водою яка знаходиться в міжклітинниках і в вільному просторі клітин.
Через зневоднення змінюється не тільки інтенсивність фотосинтезу, але і якісний склад його продуктів: менше синтезується сахарози, малату і інших органічних кислот, більше – глюкози, фруктози, аланіну та інших амінокислот.
Зниження оводненості тканин впливає на інтенсивність фотосинтезу опосередковано. По-перше, в умовах водного дефіциту зменшується фотосинтезуюча поверхня: листки повільніше ростуть, нижні листки відмирають, а у деяких рослин згортаються в трубку для зменшення транспірації. По-друге, збільшення в’язкості цитоплазми впливає на швидкість внутрішньоклітинного транспорту речовин і швидкість хімічних реакцій.
Вплив мінерального живлення. Азот і магній входять до складу пігментів; фосфор – до складу АТФ; азот, фосфор, кальцій, магній – в мембрани хлоропластів. Азот, калій, залізо, кобальт, мідь, марганець, хлор, кальцій регулюють активність ферментів; магній, азот, сірка – структуру хлоропластів.
Металопротеїди є переносниками електронів, що містять залізо (цитохроми, фередоксин), мідь (пластоціанін), марганець (комплекс, розкладаючий воду). Дефіцит цих елементів викликає різкі зміни в швидкості транспорту електронів, відновлення НАДФ.
Залізо необхідно для синтезу хлорофілу. Його дефіцит порушує синтез і функціонування пігментних систем, транспорт електронів.
Азот. Його дефіцит призводить до зменшення розмірів хлоропластів в 1,5-2 рази, листки стають дрібніше, продихи менш рухомі.
Магній. При його нестачі руйнуються тилакоїди строми і гран, мало утворюється хлорофілу. В результаті рослини не можуть поглинати сильне світло. Одночасно не¬стача азоту і заліза призводить до хлорозу — молоді листки стають блідо-жовті, поглинання вуглекислого газу знижується в 3 рази.
Сірка. Дефіцит порушує утворення тилакоїдів строми.
Фосфор необхідний для процесів фотофосфорилювання. В умовах тривалого фосфорного дефіциту знижується синтез крохмалю і збільшується синтез органічних кислот і амінокислот, зменшується активність РуБФК/О і збільшується активність ФЕП-карбоксилази.
Калій. При дефіциті порушуються грани хлоропластів, продихи погано відкриваються і закриваються, порушується водний режим.
Взаємодія факторів при фотосинтезі. Швидкість темнової фази фотосинтезу (С3-цикл) визначається активністю двох ключових реакцій циклу: реакцією карбоксилювання, швидкість якої регулюється концентрацією С02, і реакцією відновлення ФГК в ФГА, швидкість якої залежить від вмісту АТФ і НАДФ·Н2. Концентрація С02 в хлоропласті залежить від його концентрації в атмосфері та від опору його дифузії, а кількість АТФ і НАДФ·Н2 – від транспорту електронів, що обумовлений світлом. Крім того, активність всіх ферментативних реакцій залежить від температури.
Інтенсивність світла і концентрація СО2. При низькій концентрації вуглекислого газу інтенсивність фотосинтезу не залежить від освітленості, світлове насичення в таких умовах настає раніше. При низькій інтенсив¬ності світла фотосинтез не залежить від концентрації СО2. Чим менша освітленість, тим швидше настає вуглекислотне насичення. Світлова компенсаційна точка настає раніше при більш високій концентрації СО2, тому що в цих умовах збільшується швидкість фотосинтезу. Освітленість або концентрація СО2 можуть обмежувати фотосинтез.
Температура. При низьких інтенсивностях світла температура не впливає на інтенсивність фотосинтезу. При великій інтенсивності світла більш високі температури збільшують інтенсивність фотосинтезу, якщо достатньо СО2. Якщо СО2 мало, то збільшення температури не впливає на інтенсивність фотосинтезу при збільшенні освітленості.
При підвищенні температури компенсаційна точка наступа¬є при більшій освітленості, тому, що в цих умовах дихання активується сильніше, ніж фотосинтез. Зниження температури призводить до більш раннього досягнення як світлового, так і вуглекислотного компенсаційних пунктів.
Зазвичай один з факторів, який знаходиться в мінімумі, обмежує вплив інших факторів. Ця залежність отримала назву закону обмежуючих факторів. В 1905 р. Ф. Блекман (Англія) на основі цього зробив висновок, що фотосинтез складається з двох фаз: залежної від світла (світлової фази) і залежної від концентрації вуглекислого газу і температури (темнової фази). При незначній освітленості фо-тосинтез зростає пропорційно інтенсивності світла і майже не залежить від факторів, що впливають на темнові реакції фотосинтезу. При невеликій освітленості темнова фаза проходить повільно.
Денний хід фотосинтезу – зміни його інтенсивності протягом дня, обумовлені впливом різних зовнішніх факторів і їх взаємодії.
В помірному кліматі при достатньому забезпеченні водою хід фотосинтезу загалом відповідає зміні освітленості. Починаючи з сходом сонця, фотосинтез досягає максимуму в денні години, а потім поступово знижується до вечора, припиняючись з заходом сонця. В жаркому кліматі в денні години, коли через високу температуру рослина втрачає багато води, інтенсивність фотосинтезу знижується. Вона знижується тим сильніше, чим вище температура, і іноді опиняється нижче компенсаційної точки. До вечора при зниженні температури і деякому підвищенні оводненості тканин інтенсивність фотосинтезу зростає. В цих умовах крива денного ходу фотосинтезу двовершинна, та має два максимуми: більш високий вранці і менш високий ближче до вечора. Такі двовершинні криві характерні і для денного ходу фотосинтезу рослин помірної зони в найбільш жаркі дні.
В жаркі дні перегрів листка сонячними променями, знижує фотосинтез і одночасно підвищує інтенсивність дихання, при цьому листки виділяють СО2. В тропічних країнах у деяких культур (кофе, цукрова тростина) продуктивність фотосинтезу більша в похмурі дні.
На денний хід фотосинтезу впливають умови попередніх днів. Протягом декількох діб може зберігатись попередня ритмічність фотосинтезу.
В усіх кліматичних зонах є види з високою і низкою інтенсивністю фотосинтезу. В залежності від виду ця величина може змінюватись в 20-25 рази. Чим гірші умови існування, тим вище швидкість фотосинтезу, але тим менше тривалість активної фотосинтетичної роботи листка на добу.
Величина інтенсивності фотосинтезу відображає напруженість зовнішніх факторів на даний момент, є результатом стану рослини в попередні дні (післядія) і в деякому ступені передує подіям, які можуть виникнути в майбутньому. В цьому полягають причини різноманітних денних коливань швидкості фотосинтезу в природі.
Вплив внутрішніх факторів. Залежність фотосинтезу від генетичних особливостей. Інтенсивність фотосинтезу відрізняється у рослин різних систематичних груп і життєвих форм. У трав вона в основному більша, ніж у дерев.
Величина інтенсивності фотосинтезу залежить і від типу відновлення СО2. У С3-рослин вона дорівнює 20-40, у С4-рослин – 50-80 і навіть 100 мг СО2/дм2∙год, у рослин типу САМ – 3-20 СО2/дм2∙год. Ці показники отримані в штучних умовах, що жорстко контролюються, в природних умовах різниці менше.
В близькоспоріднених групах рослин швидкість фотосинтезу варіює мало. Тільки в несприятливих умовах, наприклад, під час посухи, виявляються відміни: більш посухостійкі види і сорти зберігають більш високу інтенсивність фотосинтезу.
Фотосинтетичний апарат дуже стійкий, він працює у різ¬них видів приблизно з однаковою швидкістю. Тому, що біохімічні процеси в світі тотожні.
Врожайність рослин, крім ін¬тенсивності фотосинтезу, залежить зокрема від розташування листків. Всі листки у культурних сортів менше затіняють один одного і тому поглинають більше світла. В результаті фотосинтез проходить інтенсивно не тільки в верхніх листках, але і в нижніх, і в рослині накопичується більше органічних речовин.
Інтенсивність фотосинтезу підпорядковується і ендогенним ритмам. У різних видів різна ритмічність фотосинтезу. Наприклад, при безперервному освітленні рослин квасолі інтенсивність фотосинтезу в період, що співпадає з астрономічною ніччю, менше, ніж в астрономічний день.
Залежність фотосинтезу від віку рослини. У всіх рослин на початку життєвого циклу, коли формується фотосинтетичний апарат, інтенсивність фотосинтезу зростає, а досягнувши максимальної величини, повільно зменшується.
Максимальна фотосинтетична активність листка співпадає з завершенням його формування. Після цього інтенсивність фотосинтезу деякий час не змінюється, а згодом на початку старіння листка знижується.
Інтенсивність фотосинтезу залежить від структури хлоропластів. При їх старінні руйнуються тилакоїди. Світлова фаза відбувається гірше з збільшенням віку хлоропластів. В умовах достатньої кількості води і азоту, які затримують старіння хлоропластів швидкість фотосинтезу з віком знижується повільніше.
Зменшення інтенсивності фотосинтезу з віком відбувається, навіть якщо збільшується концентрація хлорофілу. Тому, що інтенсивність фотосинтезу визначається не концентрацією хлорофілу, а структурою хлоропластів.
Швидкість фотосинтезу як хімічного процесу залежить від кількості продуктів реакції. Частина асимілятів перетворюється в первинний крохмаль в хлоропластах й виводиться з хімічних реакцій. Крохмаль вночі може перетворитись знову в сахарозу або в сорбіт і транспортуватись в інші органи. Але якщо накопичення асимілятів продовжується, то інтенсивність фотосинтезу зменшується.
|
|
Дата добавления: 2014-01-07; Просмотров: 16465; Нарушение авторских прав?; Мы поможем в написании вашей работы!