КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Хімічна природа ферментів
|
|
|
|
Відомо, що ферменти мають білкову природу. Як і білки, їх ділять на прості і складні. Прості ферменти є однокомпонентними, а складні – дво- і багатокомпонентними.
Для ферментів-білків характерна будова, типова для цього класу сполук. Для ряду ферментів була визначена первинна, вторинна, третинна і четвертна структура молекули. Так, встановлена послідовність амінокислотних залишків в молекулі панкреатичної РНК-ази, лізоциму, хімотрипсина, аспартатамінотрансферази й ін. У більшості ферментів молекула складається з одного поліпептидного ланцюга, що має вигляд a- і b-спіралей. Вивчена третинна і четвертна структури ферментів. Третинна і четвертна структури ферменту визначають характер його функцій. Так, якщо молекула має 12 субодиниць, то фермент здійснює реакцію декарбоксилування (піруватдекарбоксилаза), шість субодиниць – фермент каталізує окисно-відновні перетворення пірувата та ін.
Молекули ферментів зазвичай мають великі розміри і складну просторову конфігурацію. Кожна частина молекули ферменту строго спеціалізована. Так, в молекулі ферменту є активний центр. Він є динамічним утворенням, що містить у собі оптимальне розміщення різних функціональних груп і угрупувань, які забезпечують акт каталізу. Молекули трипсину і карбоксипептидази мають по одному активному центру, гліцеральдегід-фосфат-дегідрогенази – 2, уреази – 3 – 4, холінестерази – 20 – 100. В активному каталітичному центрі виділяються спеціалізовані ділянки, що відповідають за зв’язок з субстратом – контактна, або „якірна” ділянка (рис. 5).
Периферійні частини молекули ферменту беруть участь в підтримці і формуванні просторової конфігурації активного центру, визначають внутрішньоклітинну локалізацію ферменту і його активних центрів, взаємостосунки з клітинними мембранами і органоїдами клітини та ін. У молекулі ферменту є ділянки, з якими можуть взаємодіяти інші речовини – алостеричні активатори або інгібітори, що змінюють просторову конфігурацію і каталітичну активність всього ферменту і його активних центрів.
Активний центр ферментів. Вивчення специфічності і каталітичної дії ферментів дало можливість встановити, що субстрат зв’язується не з усією молекулою ферменту, а з окремою її ділянкою, яка називається активним центром (рис. 5). Оскільки активний центр визначає специфічність і каталітичну активність ферменту, то він повинен складатися із структури певного ступеня складності, яка може зближуватися і взаємодіяти з молекулою субстрату.
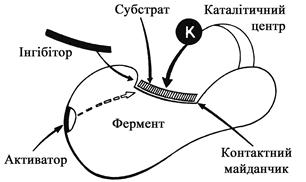
Рис. 5. Схема молекули ферменту (за О.С. Хохловим і Ю.О. Овчинниковим)
У складних ферментах (протеїдах) активний центр утворений кофактором і залишками амінокислот; у простих ферментах (протеїнах) активний центр представлений певною комбінацією залишків амінокислот, які розміщені на відповідній ділянці молекули ферменту. До складу активного центру більшості простих ферментів входять залишки амінокислот цистеїна, серина, аргініна, аспарагінової і глутамінової кислот, гістидина, тирозина і триптофана. Серед них важливе значення мають групи HS (цистину), групи OH (серину), імідазольне кільце гістидину. Певну роль відіграють функціональні групи інших залишків амінокислот, зокрема карбоксильні групи аспарагінової і глутамінової кислот та індольна група триптофану. При цьому необхідно зауважити, що такі функціональні групи розміщені в поліпептидних ланцюгах ферменту на різній відстані одна від одної. Їх зближення з утворенням активного центру відбувається тільки внаслідок формування властивої для ферменту вторинної і третинної структур. Отже, активний центр ферменту складається із функціональних груп, які відповідно зорієнтовані у просторі. Порушення третинної і вторинної структур призводить до зниження або повної втрати активності ферменту. Це можна спостерігати при дії на ферменти високої температури, тобто коли відбувається їх денатурація, яка супроводжується деформацією структури білка.
Деякі ферменти мають два і більше активних центри. В активному центрі розрізняють так звану каталітичну ділянку, яка безпосередньо взаємодіє з субстратом, і контактну (якірну) ділянку, яка зумовлює спорідненість до субстрату, формування його комплексу з ферментом.
Такий поділ активного центру ферменту є досить умовним, оскільки взаємодія в контактній ділянці фермент-субстратних сполук значною мірою впливає на швидкість перетворень, які відбуваються на каталітичній ділянці ферменту. Умовний характер має також і саме відділення активного центру від інших частин ферменту, оскільки вони мають важливе значення в утворенні його нативної структури і впливають на реакційну здатність функціональних груп активного центру. Отже, за каталітичну активність ферменту відповідає вся структура його молекули.
В ході каталітичного акту відбувається своєрідна індукційна „підгонка” субстрата (іноді і інгібітору) до активного центру (рис. 6).
Крім активного центру в молекулі ферменту може бути також алостеричний центр (центри) (від грец. алос – другий, інший і стереос – просторовий, структурний). Алостеричний центр – це ділянка молекули ферменту, яка в результаті приєднання до неї низькомолекулярної сполуки зумовлює зміну просторової (третинної), а іноді і четвертинної структури ферменту. Це, в свою чергу, призводить до зміни конфігурації активного центру, що зумовлює зміну каталітичної активності, тобто підвищує або знижує каталітичну активність. Цей процес є основою так званої алостеричної регуляції ферментативної активності. Ферменти, активність яких контролюється станом активного та алостеричного центрів, називаються алостеричними ферментами. Характерною особливістю цих ферментів є наявність у молекулі олігомерного ферменту декількох активних і алостеричних центрів. Функціональна активність цих центрів тісно взаємозв’язана між собою і структурою усієї молекули ферменту.
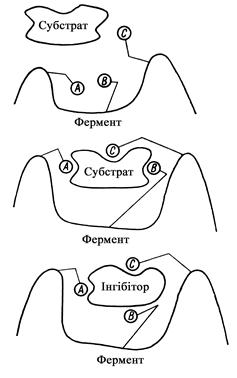
Рис. 6. Схема індукованої „підгонки” ферменту і субстрату, ферменту й інгібітору
Прості ферменти. Молекули простих ферментів при гідролізі розщеплюються до амінокислот. За хімічними властивостями прості ферменти відносяться до альбумінів (естераза печінки), глобулінів (трипсин і уреаза) та до інших груп простих білків. Простими ферментами є гідролази харчового каналу (трипсин, уреаза, РНК-аза, ДНК-аза, хімотрипсин), лізоцим яйця, алкогольдегідрогеназа дріжджів і печінки та ін. У простих ферментів активний центр зазвичай формується із залишків гістидина, серина, аргініна, триптофана, цистеїна, аспарагінової і глутамінової кислот, тирозина. Так, в активному центрі РНК-ази розміщені два залишки гістидина, залишок аспарагіна і лізину. При формуванні третинної структури ферменту в його молекулі утворюється своєрідна „кишеня” поліпептидного ланцюга, в якій і відбуваються каталітичні перетворення субстрата (рис. 5).
Складні ферменти. Більшість ферментів є складними білками. При діалізі білковий компонент (апофермент) залишається в діалізаторі, небілковий проходить через напівпроникну перегородку. За присутності небілкових компонентів (кофакторів) виявляється каталітична дія багатьох ферментів. M. Діксон і Е. Уебб всі кофактори ферментів розділили на три види: коферменти, простетичні групи і активатори. Ні кофактор, ні апофермент не можуть самостійно повноцінно здійснювати каталітичний акт. Так, при з’єднанні заліза з молекулою порфірина утворюється гем, його каталітична активність щодо розкладання пероксида водню зростає в 1000 разів, а після приєднання гема до специфічного білкового компоненту утворюється каталаза, каталітична активність її в 100 000 000 разів перевищує активність кофактора. Кофактор в середньому складає близько 1 % загальної маси ферменту.
Зв’язок між апоферментом і кофактором в молекулі різних ферментів неоднаковий. Часто кофактор неміцно зв’язаний з апоферментом і з’єднується з ним під час каталітичного акту. В цьому випадку кофактор є типовим коферментом (див. вище). Прикладом таких ферментів є дегідрогенази. В багатьох ферментах кофактор міцно і постійно зв’язаний з апоферментом – простетична група, наприклад каталаза і пероксидаза, які містять гем. До активаторів ферментів відносять такі кофактори, які переводять фермент в активний стан, але самі не беруть участь в каталітичному акті. До них відносяться багато неорганічних іонів. До ферментів такого роду можна віднести карбоксипептидазу, карбоангідразу та ін. Їх часто називають металоферментами.
Розподіл кофакторів на три види умовний, оскільки дуже часто важко провести чітку грань між коферментом і простетичною групою, між іонами металів що входять до складу молекули та іонами, що активують каталітичний процес.
Функції кофакторів різноманітні. Перш за все, вони є основою для формування активного центру і здійснення контакту молекули ферменту з молекулами субстрата. Вони беруть участь у перенесенні атомних груп, атомів, протонів і електронів у ході каталітичного процесу. Деякі з них виконують зв’язуючі функції між різнорідними ферментами, забезпечуючи узгодженість їх дії. Каталіз здійснює вся молекула ферменту. Її білкова частина обумовлює швидкість реакції і специфічність дії. Один і той же кофактор може бути складовою частиною багатьох ферментів, різних за своєю будовою і значенням. Так, фосфопіридоксаль є кофактором понад 30-ти ферментів, що здійснюють карбоксилування, транс- і переамінування.
В молекулах складних ферментів в утворенні активного центру бере участь відповідний кофактор і деякі прилеглі до нього білкові функціональні групи: SH-група цистеїна, ОН-група серина, імідазольне кільце гістидина, СООН-групи аспарагінової і глутамінової кислот, індольна група триптофана та ін.
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 767; Нарушение авторских прав?; Мы поможем в написании вашей работы!