КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Функции лизосом
|
|
|
|
Пл
Основные функции поверхностного аппарата
1. Трансмембранный транспорт. Представлен тремя категориями процессов:
а) диффузия;
б) пассивный транспорт или облегченная диффузия;
в) активный транспорт или избирательный (при участии АТФ и ферментов).
2. Рецепторная функция.
3. Транспорт в мембранной упаковке. Делится на экзоцитоз и эндоцитоз (фагоцитоз, пиноцитоз) в зависимости от того, в каком направлении переносятся вещества (в клетку или из клетки).
ЦИТОПЛАЗМАТИЧЕСКИЙ МАТРИКС. Представляет собой основную и наиболее важную часть клетки, ее истинную внутреннюю среду. Компоненты цитоплазматического матрикса осуществляют процессы биосинтеза в клетке и содержат ферменты, необходимые для продуцирования энергии, главным образом за счет анаэробного гликолиза.
Основные с в о и с т в а цитоплазматического матрикса:
1. С матриксом связаны коллоидные свойства клетки. Вместе с внутриклеточными мембранами вакуолярной системы его можно рассматривать как высокогетерогенную или многофазную к о л л о и д н у ю с и с т е м у.
2. Цитоплазматический матрикс обеспечивает изменение вязкости цитоплазмы различных клеток, которая возникает под действием внешних и внутренних факторов.
3. Цитоплазматический матрикс ответственен за циклоз, амебовидное движение, деление клетки и движение пигмента в хроматофорах.
4. Цитоплазматический матрикс определяет полярность расположения внутриклеточных компонентов.
5. Цитоплазматический матрикс обеспечивает механические свойства клеток, такие, как эластичность, способность к слиянию, ригидность.
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС). ЭПС состоит из трех морфологических компонентов: канальцев, микровакуолей и крупных цистерн.
|
|
|
Существуют две разновидности ПС:
а) шероховатая или гранулярная, когда цистерны и канальцы связаны с рибосомами.
б) гладкая или агранулярная, когда связь с рибосомами отсутствует.
Функции ЭПС
1. Обеспечивает синтез белка.
2. Обеспечивает активный транспорт различных соединений по внутримембранной фазе.
3. Синтез мембранных липидов.
4. Транспорт и накопление ионов в клетке, а также резервуар ионов кальция.
5. Синтез предшественников стероидных гормонов и других специфических соединений.
6. Детоксикация вредных продуктов метаболизма (особенно в гепатоцитах позвоночных).
7. Синтез протеолитических ферментов.
8. Первичный синтез секрета.
КОМПЛЕКС ГОЛЬДЖИ (КГ). КГ состоит из трех морфологических компонентов: уплощенные цистерны, микровакуоли и крупные цистерны (рис. 2).
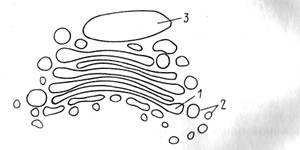
Рис. 2. Строение аппарата Гольджи: 1—уплощенные цистерны, 2 — микровакуоли, 3 — крупные цистерны
В растительных клетках цистерны отсутствуют. Такой КГ называют диктиосомы.
Особенность строения КГ
На своих мембранах КГ никогда не имеет рибосом.
Функции КГ
1. Формирование первичных лизосом с их своеобразными мембранами и сложной структурной организацией гидролаз.
2. Формирование особых структур — пероксисом или микротелец— пузырьков, ограниченных одинарной мембраной и содержащих каталазу, оксидазу Д-АМИНОКИСЛОТ, уротоксидазу и некоторые другие окислительные ферменты.
3. Сборка и «рост» мембран, которые затем окружают накапливающиеся продукты секреции, после чего они освобождаются из органеллы.
4. Обезвоживание, накопление, упаковка и транспорт продуктов секреции.
5. Синтез структурных компонентов клетки, таких, как, например, коллаген — компонент соединительной ткани.
6. Участвуют в синтезе желтка яйцеклеток и синтезе полисахароидов.
МИТОХОНДРИИ. Митохондрии ограничены двумя мембранами. Внешняя митохондриальная мембрана отделяет ее от гиалоплазмы. Обычно она имеет развитые контуры, не образует впячивания или складки. Внутренняя мембрана ограничивает собственно внутреннее содержимое митохондрий, ее матрикс. Характерной чертой внутренней мембраны митохондрий являются складки, которые имеют вид плоских гребней или крист. Как межмембранное пространство, так и полость матрикса заполнены содержимым гомогенного строения. В матриксе иногда выявляются тонкие нити и гранулы (рис. 6).
|
|
|

Рис.3. Строение митохондрии: 1 — наружная мембрана, 2— внутренняя мембрана, 3 — впячивания внутренней мембраны, 4 — места впячивания крист с поверхности внутренней мембраны
Функции митохондрий
1. Осуществляют синтез АТФ.
2. Участвуют в углеводном и азотистом обмене.
а). На наружной мембране и в окружающей ее гиалоплазме идут процессы анаэробного окисления (гликолиз).
б). На внутренней мембране митохондрий идут процессы, связанные с окислительным циклом трикарбоновых кислот и дыхательной цепью переноса электронов.
3. Осуществляют синтез белка. Митохондрии обладают полной системой синтеза белков, то есть имеют спою специфическую ДНК, митохондриальную ДНК
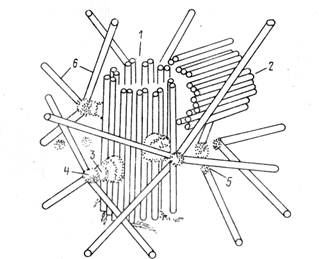
Рис. 4. Строение клеточного центра: 1—материнская центриоль, 2 — дочерние центриоли, 3 — ножка сателлита, 4 — головка сателлита, 5 — фокусы схождения микротрубочек, 6 — микротрубочки
КЛЕТОЧНЫЙ ЦЕНТР (диплосома) и центросферы носит название клеточного центра. Основу строения центриолей составляют расположенные по окружности девять групп микротрубочек (по три микротрубочки в группе), образующих таким образом полый цилиндр. Система микротрубочек центриоли описывается формулой (9+0), подчеркивая отсутствие микротрубочек в ее центральной части. Вокруг каждой центриоли расположен бесструктурный или тонковолокнистый матрикс(рис. 4).
Центриоли характерны и обязательны для клеток животных, их нет у высших растений, у низших грибов и некоторых простейших.
Строение и активность центриолей меняется в зависимости от периода клеточного цикла (интерфаза, митоз). Начиная с профазы и кончая телофазой, центриоли имеют сходное строение. В это время происходит формирование веретена митотического аппарата, состоящего из микротрубочек. То есть клеточный центр является местом полимеризации микротрубочек (рис. 8).
|
|
|
В предсинтетический период активность клеточного центра связана с образованием цитоплазматических микротрубочек (цитоскелета).
В синтетический период, помимо образования микротрубочек цитоскелета, может идти образование ресничек или жгутиков. Базальные тельца этих структур кроме периферических групп микротрубочек имеют центральную группу микротрубочек. Система микротрубочек реснички описывают как (9+1) в отличие от (9+0) системы центриолей.
При наступлении синтетического периода происходит удвоение числа центриолей. В постсинтетический период микротрубочки цитоскелета исчезают и вокруг центриолей (дипло-сом) начинают отрастать митотические трубочки.
Таким образом, функции клеточного центра заключаются:
а) в образовании цитоплазматических микротрубочек;
б) в построении веретена деления;
в) в образовании ресничек и жгутиков.
РИБОСОМЫ. Это сложные рибонуклеопротеиды (РИМ), и состав которых входят белки и молекулы РНК, примерно и равных весовых соотношениях. Работающая рибосома состоит из двух неравных субъединиц: большой и малой.
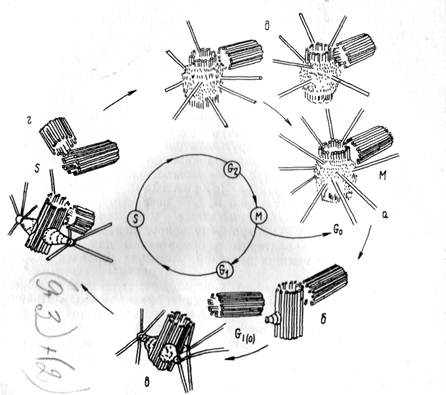
Рис. 5. Центриолярный цикл: а— диплосома во время митоза, б —центриоль
в начале G, — периода, в —центриоль в G, — периоде, г — центриоли в
S — периоде, удвоение центриолей, д — центриоли в G2 — периоде
субъединиц весьма сложна. Малая субъединица изогнута в виде телефонной трубки, а большая напоминает ковш.
Рибосомы присутствуют и в прокариотах, и в эукариотных клетках. В клетках эукариот существует две разновидности рибосом: рибосомы собственно цитоплазмы и рибосомы, локализованные в таких клеточных органоидах, как митохондрии и хлоропласты.
Рибосомы эукариот имеют коэффициент седиментации 80S коэффициент седиментации характеризует скорость осаждения частицы при ультрацентрифугировании), а прокариот — 70S. Рибосомы митохондрий и хлоропластов в общем тоже относятся к классу 70S.
|
|
|
В состав цитоплазматических рибосом эукариотных клеток входят всего четыре молекулы РНК (из них три в большой субъединице), а в рибосомах прокариот присутствуют три молекулы РНК (из них две в большой субъединице).
Белковый состав рибосом очень неоднороден. Белки большой и малой субъединиц рибосом резко отличаются по аминокислотному составу и молекулярной массе.
При укладке тяжа РНП в СУБЪЕДИНИЦЫ рибосом образуются белковые активные центры. На малой субъединице имеются два активных центра:
а) и-РНК — связывающий участок;
б) участок, удерживающий амнноацил-т-РНК. На большой субъединице различают:
а) аминоацильный центр;
б) пептидный центр.
Между этими участками выделяют еще один, частично перекрывающийся с ним, - дипептидилтрансферазный центр, который катализирует образование пептидных связей.
Рибосомы — это фабрики биосинтеза.
ЛИЗОСОМЫ. Состоят из очень пестрого класса пузырьков размером 0,2—0,8 мкм, ограниченных одиночной мембраной. Каждая лизосома содержит разнообразные ферменты, способные расщеплять биологические продукты в слабокислой среде. Содержащиеся в лизосоме ферменты окружены мембраной и благодаря этому разобщены с субстратом, находящимся в клетке.
Основные протеолитические ферменты лизосом:
1. Кислая РНКаза и кислая ДНКаза расщепляют нуклеиновые кислоты.
2. Кислая фосфотаза — фосфорные эфиры.
3. Фосфопротеидфосфотаза — фосфопротеиды и пирофосфа-ты.
4. Катепсин и коллагеназа — белки.
5. Альфа-глюкозидаза — полисахариды.
6. Бетта-глюкоронидаза — мукополисахариды.
7. Арилсульфатаза — эфиры серной кислоты.
Среди различных по морфологии лизосомных частиц можно выделить по крайней мере четыре типа:
1. Первичные лизосомы формируются из вакуолей периферической зоны аппарата Гольджи и заполнены протеолитическими ферментами в неактивной форме, ранее синтезированными на гранулярной эндоплазматической сети (рис. 9).
2. Вторичные лизосомы или внутриклеточные пищеварительные вакуоли по своей морфологии представляют собой первичную лизосому, слившуюся с фагоцитарными или пиноцитозными вакуолями. От типа поглощенных веществ или частичек зависит размер и внутренняя структура таких лизосом. Судьба поглощенных биогенных веществ, попавших и состав лизосомы, заключается в их расщеплении гидролазами до мономеров и в транспорте этих мономеров через мембрану лизосомы в состав цитоплазмы, где они включаются в различные синтетические и обменные процессы.

Рис. 6. 1,12 —крупные и мелкие частицы в среде, окружающей клетку, 3,4 — захват частиц путем фагоцитоза и возникновение лизосом (первичная лизосома), 7 — фаголизосома, 8 —пищеварительное тельце, 9 — остаточное тельце, 10 — выделение продуктом разложения 11 —аутолизосомы
3. Телолизосомы или остаточные тельца. Такие лизосомы возникают на вторичных, при переваривание биогенных микромолекул внутри лизосом идет не до конца. Остаточные тельца уже содержат много протеолитических ферментов, в них происходит уплотнение содержимого и его отложение в виде сложных структур. Судьба остаточных телец может быть двоякой: одни из них выбрасываются из клетки путем экзоцитоза, другие же остаются в клетках вплоть до их гибели (например, липофусциновые гранулы).
4. Аутолизосомы (аутофагосомы). По своей морфологии их относят к вторичным лизосомам, по г тем отличием, что в составе этих вакуолей встречаются фрагменты или даже целые цитоплазматические структуры. В этом случае лизосомы выполняют роль внутриклеточных чистильщиков, контролирующих дефектные структуры.
1. Гетерофагическая функция — принимают участие гидролитической обработке чужеродных веществ, поступающих в клетку при фагоцитозе и пиноцитозе (микрофаги).
2. Защитная функция — образуют мощно развитую систему лизосомных аппаратов в свободных клеточных элементах (макрофаги), которые реализуют функции адаптивного иммунитета.
3. Участвуют во внутриклеточном пищеварении— главном способом питания одноклеточных эукариотных клеток.
4. Выполняют функцию эндогенного питания в условиях голодания многоклеточных организмов, идет переваривание при помощи лизосом части низкоэнергетических структур и усвоение образующихся низкомолекулярных соединений на нужды энергетического обмена.
5. Специфическая аутофагия — при необходимости и утилизация избытков секрета в клетке, а также регулируемая деятельность лизосом в эмбриогенезе, в процессах и при дифференцировке клеток.
6. Полная клеточная аутофагия при нарушении метаболизма изоляции внутрилизосомальных протеолитических ферментов
МИКРОТЕЛЬЦА ИЛИ ПЕРЕКСИСОМЫ. Это небольшие вакуоли (0,3—1,5 мкм) с одинарной мембраной, ограничивающие гранулярный матрикс. Микротельца, подобно лизосомам, происходят из эндоплазматической сети клетки. В них обнаруживаются ферменты, связанные с метаболизмом перекиси водорода (оксидазы, уракситодазы и др.).
Функции микротелец:
1) защитная— нейтрализуют перекись, которая является токсическим веществом для клеток;
2) образуют депо ряда ферментов, которые играют важную роль при превращении жиров в углеводы.
ЯДЕРНЫЙ АППАРАТ. Имеет различную организацию в интерфазе — это рабочая фаза клеточного ядра или время, когда хромосомы функционируют.
Строение ядра в интерфазе (рис. 7).
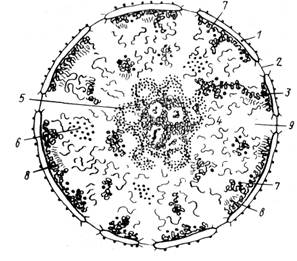
Рис. 7. Интерфазное ядро: 1 — ядерная оболочка (две мембраны, внешняя и внутренняя, и между ними перинуклеарное пространство, 2 — ядерная пора, 3 — конденсированный хроматин, 4 —Диффузный хроматин 5 —ядрышко, 6 — интерхроматиновые гранулы, 7 — перихроматиновые гранулы, 8 хроматиновые фибриллы, 9 — кариоплазма, ядерный сок
Как правило, в интерфазном ядре видны следующие структуры:
1. Ядерная оболочка обеспечивает компактизацию ядерного аппарата и формирует связь с мембранными компонентами цитоплазмы (ЭПС и КГ). Ядерная оболочка построена двумя замкнутыми мембранами, разделенными перинуклеарным пространством. Во многих местах ядерной оболочки образуются округлые перфорации, заполненные сложной структурой — комплексом ядерных пор.
2. Кариоплазма или нуклеоплазма, или ядерный сок заполняет все внутреннее пространство ядра между его компонентами.
3. Диффузный хроматин, представлен различными гранулярными и фибриллярными образованиями.
Хроматин в сущности форма существования хромосом в интерфазном ядре до начала их организации и уплотнения перед следующим клеточным делением.
4. Г л ы б к и хроматина.
Функциональное значение их пока неизвестно.
5. Ядрышко. Тельца, обычно округлой формы, сильно преломляющие свет. Ядрышко — не самостоятельная структура или органоид — оно производное хромосомы, один из ее локусов, активно функционирующий в интерфазе. Ядрышко является местом образования рибосомных РНК и рибосом.
Белки хроматина
Во фракции хроматина весовые соотношения ДНК: гистоны: негистоновые белки: РНК липиды равны 1: 1: 0,2: 0,1:: 0,01.
Гистоны — основные белки, играющие роль и обеспечении не только в упаковке хромосомной ДНК, по и играют роль в регуляции транскрипции
Негистоновые белки представляют собой специфические белки — регуляторы, узнающие определенные- нуклеотидные последовательности в ДНК-РНК хроматина представлена всеми перечисленными клеточными типами.
Липиды, обнаруженные в хроматине, вероятно, выполняют структурную роль в организации хромосом.
Структурная организация хроматина Как митотическая, так и работающая интерфазная хромосома в основе своего строения имеет элементарную хромосомную фибриллу, молекулу дезоксирибонуклеопротеида (ДНП). Существует несколько степеней структурной укладки молекул ДНК, первичным из которых является хромосомная фибрилла.
Хромосомная фибрилла расщепляется на фрагменты стандартной длины (около 200 нуклеотидных пар). Каждый фрагмент, получивший название нуклеосомы, состоит из глобулы и свободного нитьевидного участка. Нить — ДНК, а глобула—«сердцевина» нуклеосомы. В состав «сердцевины» входят восемь гистонов, вокруг которых спирально (1,75 витка) уложена нить ДНК. Остальная часть ДНК (около 60 нуклеотидных пар) образует так называемый «линкер» или межиуклеосомный участок. С этим «линкерным» участком, так и с «сердцевиной» связан гистон Н,, который обеспечивает п растворе сближение нуклеосом (рис. 8).
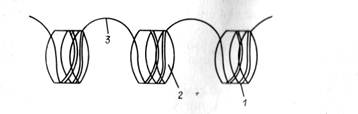
Рис. 8. Схема строения нуклеосом: 1— ДНК накрученная на сердцевину, 2 — сердцевина нуклеосомы (октомер). 3—«линкер»
Уровни компактизации ДНК (рис. 9).
Первый уровень — нуклеосомпый — образует структуру хроматиновых фибрилл в виде «бусинок на нитке», при этом происходит укорочение ДНК примерно в 7 раз. 
Рис. 9. Уровни компактизации хромосом: 1—нуклеосома, 2— нуклеомер, 3 — хромомомера 4 хромонема, 5—хромосома
Второй уровень —н у к л е о м е р н ы й, где идет объединение 8—10 нуклеосом в глобулы.
Третий уровень — хромомерный, где нуклеомерные фибриллы формируют многочисленные петли, объединенные скрепками из негистоновых белков.
Четвертый уровень - хромонемный, за счет сближения в линейном порядке хромомерных петель с образованием хромонемной нити.
Пятый уровень — хромосомный, образуется в результате спиральной укладки хромосомы (или хроматиды).

Рис. 10. Классификация хромосом: 1 — телоцентрическая (центромера отсутствует, либо расположена на конце хромосомы), 2 — акроцентрнчсская (центромера смещена к концу плеча хромосомы), 3 — субметацентрическая (центромера делит хромосому на два неравных плеча), 4 — метацентрическая (центромера делит хромосому на два равных плеча)
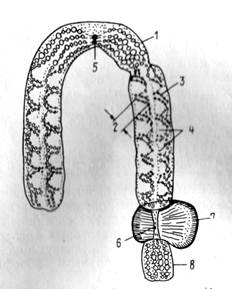
Рис. 11. Строение хромосомы (субметацентрической): 1 — пеликула, '2. —
хромонема, 3—хромомера, 4—хроматида, 5—центромера, 6 — вторичная
перетяжка, 7 — ядрышковый организатор, 8 — спутник
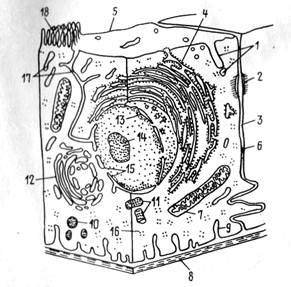
Рис. 12. Обобщенное строение клетки: 1 — пиноцитозные канальца, 2 десмосома, 3—межклеточная щель, 4 — шероховатая ЭПС, 5 —клеточная мембрана, 6 — плотный контакт, 7— митохондрия, 8— базальная мембрана, 9 — базальные лакуны, 10 — лизосомы, 11—центриоли, 12 — аппарат Голь-джи, 13 — хроматин, 14 — ядрышко, 15 — ядерная оболочка с порами, 16 — рибосомы, 17 — гладкая ЭПС, 18 — микроворсинки
ПЛАСТИДЫ. Это органеллы, теснейшим образом связанные с процессами обмена в растительных клетках. Они встречаются у всех растений, за исключением некоторых бактерий, водорослей, миксомицетов и грибов. Для этих органоидов характерно наличие пигмента (хлорофилл и каротиноиды), а также способность синтезировать и накапливать запасные вещества (крахмал, жиры и белки).
В зависимости от местоположения в растении различают три типа пластид:
1. Лейкопласты — обнаружены в клетках меристемы, эмбриональных и половых клетках, а также в неосвещенных частях растений.
2. Хромопласты - нефотосинтезирующие окрашенные пластиды, содержащие главным образом красные, оранжевые и желтые пигменты (каротиноиды). Особенно их много в зрелых плодах томатов и клетках некоторых водорослей.
3. Хлоропласты - фотосинтезирующие окрашенные пластиды, содержащие хлорофилл и отчасти каротиноиды.
Химический с о с т а в х л о р о п л а с т о в. Около 80% белка составляют нерастворимые белки, связанные с липидами (липопротеиды). Хлорофилл, как и пигменты животного организма— гемоглобин и цитохромы, представляет собой порфирин, но в его молекуле место железа занимает магний. Другие пигменты растений, относящиеся к группе каротиноидов. Хлоропласты имеют специальную нехромосомную генетическую систему (ДНК, РНК), я также содержат некоторые цитохромы, витамины К и К, атомы металлов, например, железо, медь, магний и цинк.
Строение хлоропластов (рис. 13). Этот органоид, подобно митохондриям, окружен плазматической мембраной и заполнен матриксом или стромой. В строме располагаются
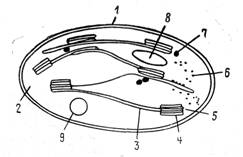
Рис. 13. Строение хлоропластов: 1—наружная мембрана, 2 — строма, 3 — ламелла между гранами, 4 — тилакоид, 5—• стопка гран, 6 — рибосомы,. 7 — капелька жира, 8 — крахмальное зерно, 9 — ДНК хлоропласта
внутренние мембраны или л а м е л ы хлоропласта, имеющие более сложное строение. Эти структуры сообщаются между собой. Многие из таких слившихся мембран уложены в стопки, или граны, соединяющиеся кое-где длинными одиночными мембранами.
Внутренняя мембранная система — это то место, где протекают световые реакции фотосинтеза. В строме проходят темновые реакции фотосинтеза, в ней находя и растворимые ферменты, в том числе ферменты цикла Кальвина, а также сахара и органические кислоты.
Функция хлоропластов. Осуществляют фотосинтез.
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 3764; Нарушение авторских прав?; Мы поможем в написании вашей работы!