КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Утворення вуглеводів при фотосинтезі
Обмін вуглеводів
Вуглеводи (моносахариди) утворюються в процесі фотосинтезу, у темновій його фазі, або, інакше, у вторинних реакціях. При синтезі вуглеводів використовуються продукти світлової фази, або первинних реакцій, – АТР і NADPH. Цикл реакцій відновлення СО2 до рівня вуглеводів, як відомо, називається циклом Кальвіна, або відновним пентозофосфатним циклом.
Цикл Кальвіна вивчають у курсі фізіології рослин, але його реакції мають бути розглянути й у курсі біохімії рослин, оскільки і вони пов'язані з первинним синтезом і перетвореннями моносахаридів.
Цикл Кальвіна виник на основі більш давнього циклу реакцій перетворення моносахаридів – окисного пентозофосфатного циклу (ПФЦ), що відбувається в процесі дихання. У циклі Кальвіна реакції йдуть у зворотному напрямку до реакцій ПФЦ. У ньому з'явилися дві реакції, що в інших метаболічних шляхах не зустрічаються, вони характерні тільки для процесу фотосинтезу. Це реакція карбоксилювання і реакція фосфорилювання рибулозо-5-фосфату.
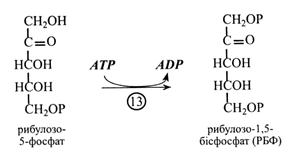
Розглянемо реакції циклу Кальвіна, яких налічується 13.
Ключова реакція циклу Кальвіна – карбоксилювання, у якій СО2 реагує з акцептором – рибулозо-1,5-бісфосфатом (РБФ).
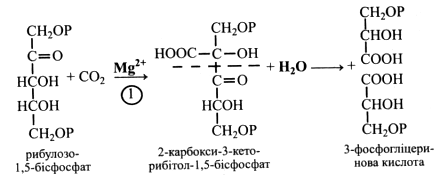
СО2 приєднується до РБФ, і утворюється проміжна сполука з шести вуглецевих атомів – цукор 2-карбокси-3-кето-рибітол-1,5-бісфосфат. Ця сполука нестійка і під дією води розпадається на дві тріоди – 2 молекули 3-фосфогліцеринової кислоти (3-ФГК).
Цю складну першу реакцію циклу каталізує фермент рибулозобісфосфат-карбоксилаза (РБФК). За вмістом він становить 50 % розчинних білків листків, а загальна кількість його у природі перевищує вміст усіх інших білків-ферментів разом узятих.
Молекулярна маса РБФК дорівнює 550000. Молекула її складається з 16 субодиниць: 8 великих (молекулярна маса 51000 – 58000) та 8 малих (молекулярна маса 12000 – 18000). У великих субодиницях присутній реакційний центр ферменту, малі – відіграють регуляторну роль.
Для протікання 1-ї реакції необхідні іони Mg2+. Вони активують РБФК і стабілізують проміжну сполуку – 2-карбокси-3-кето-рибітол-1,5-бісфосфат.
У 2-й реакції 3-ФГК під дією ферменту фосфогліцераткінази фосфорилюється за участю АТР з утворенням 1,3-ФГК.
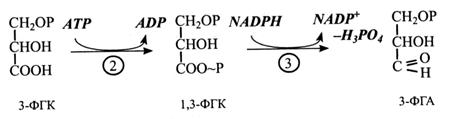
Далі 1,3-ФГК відновлюється за участю NADPH до 3-фосфогліцеринового альдегіду (3-ФГА). Реакцію каталізує тріозо-фосфатдегідрогеназа.
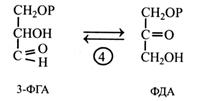
Фосфогліцериновий альдегід ізомеризується з фосфодигідроксиацетоном (ФДА). Цю реакцію каталізує фермент тріозофосфатізомераза. ФГА і ФДА з'єднуються під дією ферменту альдолази. Він каталізує подібну реакцію в ПФЦ і зворотну – в гліколізі. Цей фермент поширений у рослинах.
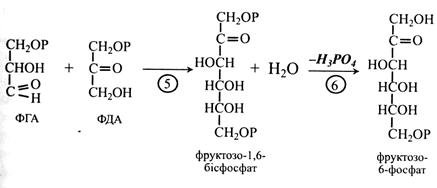
У результаті альдолазної реакції ФГА і ФДА, з'єднуючись, утворюють гексозу – фруктозо-1,6-бісфосфат. Далі в 6-й реакції від фруктозо-1,6-бісфосфату гідролітично під дією фосфатази відщеплюється один залишок неорганічного фосфату й утворюється фруктозо-6-фосфат. Частина молекул цього сахарофосфату виходить з циклу Кальвіна як його продукт, а частина залишається в циклі і бере участь у реакціях регенерації акцептора СО2 – РБФ. Встановлено, що на 1 молекулу фруктозо-6-фосфату, який виходить з циклу, припадає 5 молекул, що беруть участь у регенерації акцептора СО2.
Далі в циклі Кальвіна (7-ма реакція) фруктозо-6-фосфат взаємодіє з ФГА.
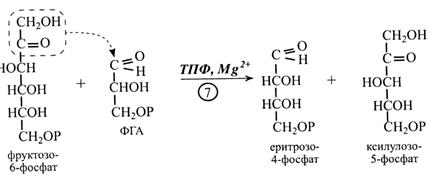
Реакцію каталізує фермент транскетолаза, коферментом якого є тіамінпірофосфат (ТПФ). Транскетолаза при участі ТПФ переносить двовуглецевий залишок з кетогрупою з молекули фруктозо-6-фосфату на ФГА. В результаті утворюються еритрозо-4-фосфат і ксилулозо-5-фосфат. Реакція активується іонами Мg2+.
У наступній (8-й) реакції еритрозо-4-фосфат і ФДА утворюють семивуглецевий сахарофосфат – седогептулозо-1,7-бісфос-фат. Реакцію каталізує альдолаза.
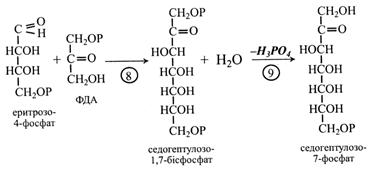
Від седогептулозо-1,7-бісфосфату під дією фосфатази відщеплюється неорганічний фосфат і утворюється седогегпулозо-7-фосфат.
У наступній реакції (10-й) знову бере участь транскетолаза, що переносить двовуглецевий залишок з молекули седогептулозо-7-фосфату на молекулу ФГА з утворенням двох пентоз – ксилулозо-5-фосфату і рибозо-5-фосфату.
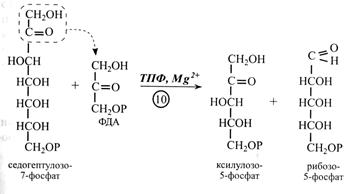
У наступних двох реакціях (11-й і 12-й) відбувається перетворення синтезованих пентоз у рибулозо-5-фосфат. Спочатку рибозо-5-фосфат ізомеризується під дією рибулозофосфат-ізомерази з рибулозо-5-фосфатом, а потім ксилулозо-5-фосфат теж перетворюється в рибулозо-5-фосфат за участю ферменту рибулозофосфат-епімерази, що змінює положення -Н та -ОН біля третього і вуглецевого атому.
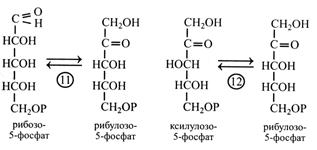
Закінчується цикл Кальвіна реакцією фосфорилювання рибулозо-5-фосфату в РБФ, тобто остаточною регенерацією акцептора СО2. Реакцію каталізує фермент фосфорибулокіназа за участю АТР.
Як зазначалося, перша й остання реакції характерні тільки для циклу Кальвіна. Решта реакцій зустрічається і в інших процесах перетворення цукрів (гліколізі, ПФЦ тощо).
Синтез, розпад і перетворення вуглеводів у рослині
Із фосфорних ефірів цукрів, що синтезуються при фотосинтезі, утворюються всі інші вуглеводи в рослині. Із хлоропластів у цитоплазму виходять різні моносахариди, що синтезуються в циклі Кальвіна. Однак найчастіше з хлоропластів виходить ФДА, що вже в цитоплазмі ізомеризується з ФГА, а потім вони з'єднуються під дією альдолази, утворюючи фруктозо-1,6-бісфосфат. Останній відщеплює Н3РО4 та утворює фруктозо-6-фосфат.

Ізомераза перетворює фруктозо-6-фосфат у глюкозо-6-фосфат, а відповідна мутаза переносить фосфатний залишок у положення 1, утворюючи глюкозо-1-фосфат. Відповідна фосфатаза гідролітично відщеплює фосфатний залишок, у результаті утворюється вільний цукор.
Важливу роль у взаємоперетворенні цукрів і біосинтезі полісахаридів відіграють нуклеозиддифосфатмоносахариди (NDP-цукри), у яких цукор з'єднаний глікозидним зв'язком із кінцевим фосфатним залишком: уридин-, цитозин-, аденозин- або гуанозиндифосфату (UDP, CDP, ADP, GDP). Із NDP зв'язуються різні моносахариди та їх уронові кислоти.
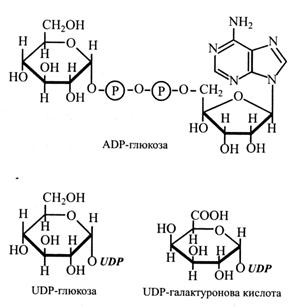
NDP-цукри утворюються з моносахарид-1-фосфатів. Наприклад:
глюкозо-1-Р + UTP «UDP-глюкоза + Н4Р2О7.
Фермент, що каталізує таку реакцію, звичайно називають просто пірофосфорилазою. Реакція утворення NDP -цукрів оборотна, але в рослинах вона спрямована, як правило, убік синтезу, тому що відбувається постійне видалення пірофосфату під дією пірофосфатази:
Н4Р2О7 + Н2О → 2Н3РО4.
Розщеплюються NDP-цyкpи фосфоролітично на NDP і цукор-1-Р:
UDP-глюкоза + Н3РO4 → UDP + глюкозо-1-Р.
NDР-цукри мають великий запас вільної енергії та беруть участь у синтезі полісахаридів, виступаючи донорами глікозильних залишків.
Біосинтез сахарози
У рослинах присутні два ферменти, що каталізують утворення сахарози: сахарозо-синтаза і сахарозофосфат-синтаза. Сахарозо-синтаза каталізує таку реакцію:
UDP-глюкоза + фруктоза → сахароза + UDP.
Вважають, що цей фермент каталізує переважно розпад сахарози, а не її синтез. Синтезується сахароза в основному за участю сахарозофосфат-синтази:
UDP-глюкоза + фруктозо-6-Р → сахарозо-6-Р + UDP;
сахарозо-6-Р + Н2O → сахароза + Н3РO4.
При цьому синтез сахарози відбувається в два етапи. Спочатку з UDP-глюкози і фруктозо-6-фосфату утворюється сахарозо-6-Р і звільняється UDP. У 2-й реакції під дією фосфатази від сахарозо-6-Р гідролітично відщеплюється Н3РO4 і утворюється вільна сахароза.
Сахароза синтезується в цитоплазмі рослинних клітин, утворення її в хлоропластах ще остаточно не доведене.
Подібно до сахарози, утворюються й інші дисахариди. Наприклад, трегалоза синтезується під дією трегалозофосфат-синтази:
UDP-глюкоза + глюкозо-6-P → трегалозо-6-P + UDP;
трегалозо-6-P + H2O → трегалоза + Н3РO4.
Розщеплюється сахароза гідролітично за допомогою сахарази, а також при обертанні реакції за участю сахарозо-синтази:
сахароза + Н2O → глюкоза + фруктоза.
Біосинтез три-, тетра- і пентасахаридів
Синтез олігосахаридів (рафінози, стахіози, вербаскози) відбувається в результаті послідовного приєднання залишків галактози. Донором галактози є галактинол. Галактинол утворюється з UDP-галактози і міоінозиту. Міоінозит – це циклічний шестиатомний спирт.
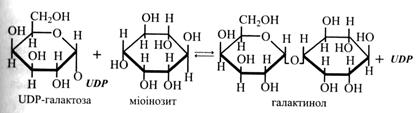
Далі залишок галактози переноситься від галактинолу спочатку на сахарозу з утворенням рафінози, потім на рафінозу з утворенням стахіози і на стахіозу з утворенням вербаскози. В усіх них реакціях виділяється вільний міоінозит:
галактинол + сахароза → рафіноза + міоінозит;
галактинол + рафіноза → стахіоза + міоінозит;
галактинол + стахіоза → вербаскоза + міоінозит.
Біосинтез вищих полісахаридів
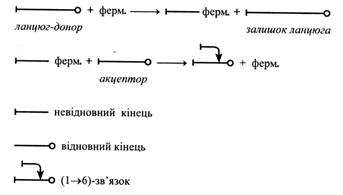
Вищі полісахариди в рослинах синтезуються шляхом реакцій трансглікозилювання, тобто переносу глікозильних залишків (залишків моносахаридів). Перенос здійснюється від численних молекул-донорів до одного кінця молекули-акцептора, яку ще називають затравкою і яка при цьому подовжується:
n глюкоза-Х + акцептор → (глюкоза)n-акцептор + nХ
донор затравка
При цьому глікозильний залишок приєднується глікозидним гідроксилом до одного з гідроксилів затравки. Численні реакції переносу каталізує один фермент, що визначає ідентичність глікозильних залишків і природу глікозидного зв'язку в даному полісахариді. Донорами глікозильних залишків частіше виступають NDP -цукри, а акцептором – частина молекули полісахариду, іноді дуже невелика, але така, що має зв'язки, характерні для макромолекули.
Біосинтез і розпад крохмалю
Крохмаль, як відомо, складається з двох полісахаридів – амілози й амілопектину. Тому його біосинтез відбувається в два етапи. Спочатку утворюється ланцюжок амілози, а потім частина її розгалужується. Інакше кажучи, спочатку синтезуються (α1→4)-зв'язки, а потім – (α1→6).
Утворення амілози каталізує фермент – крохмаль-синтаза, що знаходиться у хлоропластах і амілопластах. Донором глюкозних залишків виступає АDР-глюкоза, а акцепторами можуть бути уламки крохмальної молекули – амілози, або амілопектину. У першому випадку буде синтезуватися молекула амілози, а у другому – крохмаль-синтаза бере участь в утворенні амілопектину, та його ланцюжків з (α1→4)-зв'язками:
АDР-глюкоза + акцептор → α-1,4-глюкоза-акцептор + АDР.
Амілопектин утворюється при спільній дії крохмаль-синтази і 1,4-α-глюкан – ферменту, що розгалужує. Він переносить олігосахаридний фрагмент із невідновного кінця ланцюжка до шостого вуглецевого атома некінцевого залишку глюкози, прикріплюючи його (α1→6)-зв'язком:
Донором олігосахаридного фрагменту та його акцептором може бути як молекула амілози, так і зовнішній ланцюг амілопектину.
У розщепленні крохмалю беруть участь 6 ферментів: 5 гідролаз і фосфорилаза. Причому ці ферменти діють по-різному: одні розщеплюють (α1→4)-зв'язки, інші – (α1→6)-зв'язки, одні діють на великі молекули, інші – на їх дрібні фрагменти.
Дуже активним і поширеним у рослинах є фермент α-амілаза. З її роботи починається розщеплення крохмалю. Тільки вона діє на цілі крохмальні зерна, розщеплюючи (α1→4)-зв'язки. Після а-амілази у процес вступають інші ферменти. Кінцевими продуктами розщеплення крохмалю є глюкоза і глюкозо-1-Р.
Біосинтез і розпад целюлози
У 1964 р. Хассид із співробітниками вперше спостерігали синтез целюлозоподібної сполуки в гомогенаті з машу, а потім з гороху, кукурудзи, насіння бавовнику тощо. Донором глюкози В цих реакціях виступала GDР-глюкоза: n GDР-глюкоза + акцептор → (глюкоза)n-акцептор + nGDР.
У результаті такої реакції утворювалися не молекули целюлози, а більш короткі ланцюги полісахариду, який містить (β1→4)-зв'язки. Тоді виникло припущення, що складні комплекси ферментів, які синтезують цілі мікрофібрили целюлози і вбудовані у зовнішню поверхню плазмалеми, руйнуються при одержанні безклітинного ферментного препарату. Очевидно, синтез целюлозоподібних ланцюгів у цих умовах здійснюють окремі компоненти ферментного комплексу.
Пізніше знайшли ще одне пояснення неповного синтезу целюлозної молекули в безклітинних ферментних системах. Справа в тому, що NDР-цукри – це водорозчинні сполуки, а в мембрані, де в клітині утворюється целюлоза, середовище гідрофобне, тому і донори цукрів повинні мати гідрофобні властивості. Такими донорами в мембрані можуть бути жироподібні речовини з довгим вуглеводневим ланцюжком – поліпреноли. Вони, мабуть, приймають глюкозу від NDР-цукрів і переносять її на акцептор при синтезі целюлози в плазматичній мембрані рослинної клітини.
Розпад целюлози відбувається під дією двох гідролітичних ферментів. Целюлаза розщеплює молекули целюлози до целобіози, а потім останню гідролізує целобіаза до глюкози:

Утворення геміцелюлоз і пектинів
У геміцелюлоз і пектинів, що складають матрикс клітинної оболонки, спільні шляхи синтезу. Ферменти, що каталізують їх утворення, локалізовані в апараті Гольджі.
Взаємозв'язок шляхів біосинтезу компонентів матриксу клітинної оболонки можна побачити на наступній схемі.
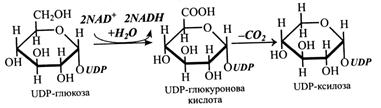
Біосинтез геміцелюлоз і пектинів починається з UDP-глюкози і UDР-галактози, які окислюються до відповідних UDР-уронових кислот, останні можуть взаємоперетворюватися під дією відповідних епімераз. Далі при декарбоксилюванні глюкуронова кислота утворює UDР-ксилозу, а галактуронова – UDР-арабінозу, які також здатні епімеризуватися.
Геміцелюлози утворюються з UDP-похідних глюкози, манози, ксилози, арабінози, галактози та уронових кислот. Для синтезу пектинів необхідні UDP-галактуронова кислота та UDP-рамноза. Крім того, для утворення розчинного пектину потрібні метальні групи, донором яких є S-аденозилметіонін (SAM). Метильні групи на полігалактуронову кислоту переносяться метилтрансферазою.
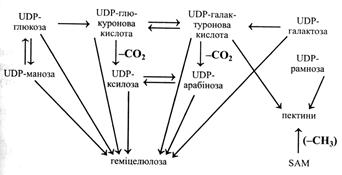
Усі реакції біосинтезу компонентів матриксу клітинної оболонки доведені дослідами in vitro. Наприклад, у достигаючому насінні кукурудзи була виявлена ферментна система, що включає ксилозу і L-арабінозу з їх UDP-похідних у ксилани. Крім того, у них були знайдені епімерази, що каталізували взаємне перетворення UDP-ксилози та UDP-арабінози.
Утворення пентоз
У рослинах існує три основних шляхи утворення пентоз – важливих для рослинного організму моносахаридів.
1. Пентози утворюються з уронових кислот. Насамперед уронові кислоти, точніше їх NDP-похідні, утворюються з NDP-цукрів шляхом окислювання останніх за участю NAD+ і Н2О. Реакцію каталізують відповідні дегідрогенази. Далі NDP-уронові кислоти декарбоксилюються з утворенням NDP-ксилози або іншої NDР-пентози.
2 Пентози виникають з альдонових кислот. Наприклад, у пентозофосфатному циклі рибулоза утворюється при окисному декарбоксилюванні фосфоглюконової кислоти, яка, у свою чергу, є продуктом окислення глюкозо-6-фосфату.
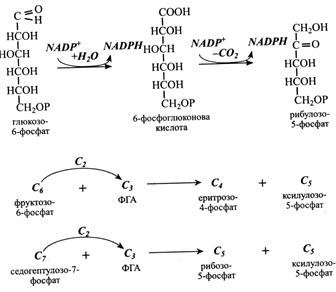
3. Пентози утворюються за участю ферменту транскетолази. Цим шляхом вони виникають у пентозофосфатному циклі й у циклі Кальвіна. При цьому фермент за участю ТПФ переносить двовуглецевий залишок з одного цукру на молекулу іншого.
Пентози, що утворилися тим чи іншим шляхом, можуть взаємоперетворюватися за допомогою відповідних ізомераз і епімераз.
Контрольні питання і завдання:
1. Дайте загальну характеристику вуглеводам. На чому базується їхня класифікація?
2. Які функції вуглеводів у рослинах?
3. Розкажіть про практичне застосування вуглеводів.
4. Як класифікують моносахариди? Дайте характеристику окремим представникам різних груп моносахаридів.
5. Охарактеризуйте олігосахариди. Як їх класифікують? Наведіть приклади основних ди-, три-, тетра- і пентасахаридів. Які їхні особливості та значення?
6. Розкажіть про солодкість цукрів, природні і синтетичні замінники цукру.
7. Дайте характеристику вищим полісахаридам. Які їхні спільні властивості?
8. Назвіть полісахариди, що входять до складу клітинної стінки; запасні полісахариди. Охарактеризуйте їхні будову, властивості, поширення.
9. Розкажіть про калозу, камеді (гумі) і слизи, їхні властивості, поширення і роль у рослині.
10. Які вищі полісахариди водоростей ви знаєте? Дайте їм характеристику. Що таке ліхенін?
11. У чому схожість і різниця між хітином й целюлозою?
12. Як утворюються вуглеводи у процесі фотосинтезу? Розкажіть про реакції циклу Кальвіна.
13. Дайте загальну характеристику обміну вуглеводів у рослинах. Яка роль NDР-цукрів у цьому процесі?
14.Розкажіть про синтез і розпад сахарози, три-, тетра-, i пентасахаридів у рослинах.
15.Які шляхи біосинтезу і розпаду крохмалю?
16.Що відомо про синтез і розпад целюлози?
17.Як відбувається синтез геміцелюлоз і пектинів?
18.Які основні шляхи утворення пентоз у рослинах?
Література:
1. Кретович В. Л. Биохимия растений. – М.: Высш. шк., 1980. – 503 с.
2. Плешков Б. П. Биохимия сельскохозяйственных растений. – М.: Колос, 1980. – 495 с.
3. Гудвин Т., Мерсер Э. Введение в биохимию растений: В 2 т. – М.: Мир, 1986.
4. Степаненко Б. Н. Химия и биохимия углеводов (моносахариды). – М.: Высш. шк., 1977. – 237 с.
5. Степаненко Б. Н. Химия и биохимия углеводов (полисахариды). – М.: Высш. шк., 1978. – 256 с.
6. Тарчевский И. А. Основы фотосинтеза. – М.: Высш. шк., 1977. –2 53 с.
|
|
Дата добавления: 2014-01-07; Просмотров: 4306; Нарушение авторских прав?; Мы поможем в написании вашей работы!