КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
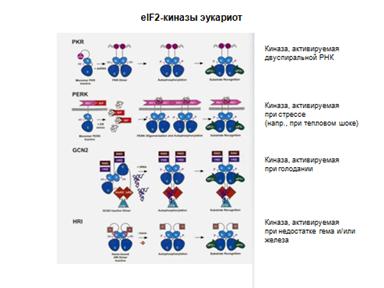
EIF2-киназы: фосфорилирование Ser51 a-субъединицы eIF2
|
|
|
|
Индукторы eIF2-киназ:
(1) Недостаток Fe3+ и/или гема
(2) Вирусная инфекция
(3) Голодание
(4) Недостаток ростовых факторов
(5) Тепловой шок
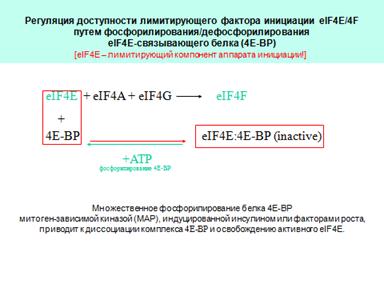
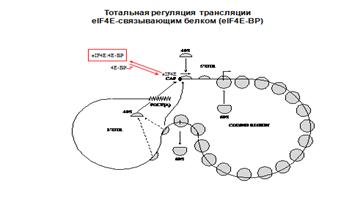
Активация синтеза белка 4Е+4А+4Г=4Ф. Есть белок 4E-ВР. При его фосфорилировании получается диссоциация 4Е и 4ЕВР.кол-во инициирующих РНК снижается.
Регуляция доступности лимитирующего фактора инициации eIF4E/4F
путем фосфорилирования/дефосфорилирования
eIF4E-связывающего белка (4E-ВР)
[eIF4E – лимитирующий компонент аппарата инициации!]
Множественное фосфорилирование белка 4Е-ВР митоген-зависимой киназой (МАР), индуцированной инсулином или факторами роста, приводит к диссоциации комплекса 4E-BP и освобождению активного eIF4Е.
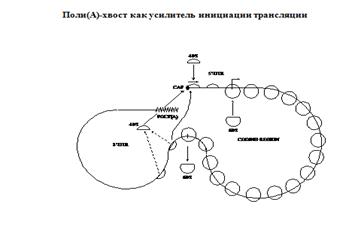
Регуляция трансляции
у эукариот:
Энхансеры трансляции находятся в 3"нетранслируемой области.
Есть 3" Экзонуклеазы.
Поли А нужен, чтобы соединится с кэпом. ПолиА связывающий белок РАВР связывается с 4Ф.

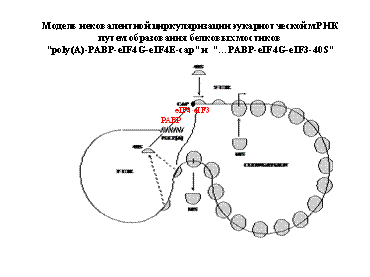
Рибосомы образуют полисому. Фотографии:

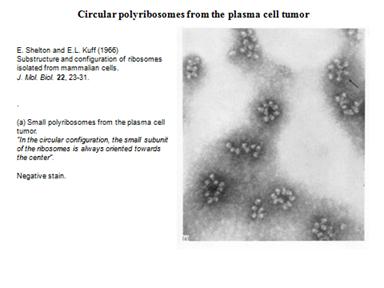
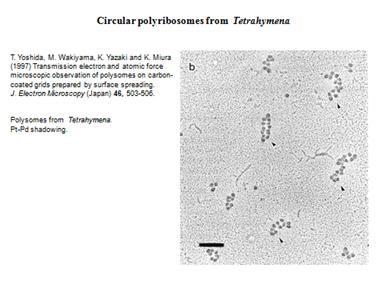
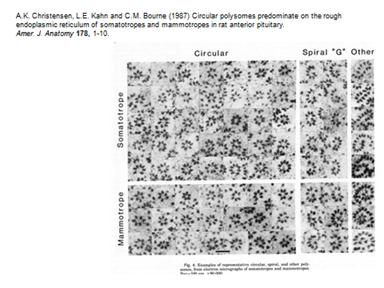
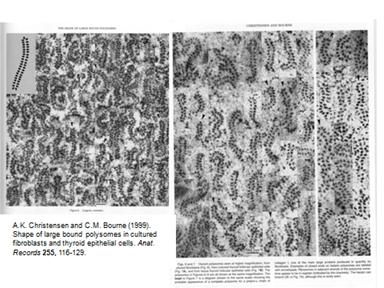
Образовываются циркулярные полисомы. Добавляют ингибитор расщепления АТФ, который ингибирует энергозависимый процесс на 5"конце. Синтез продолжался почти на прежнем уровне. Это говорит о том, что механизм трансляции идет без сканирования.
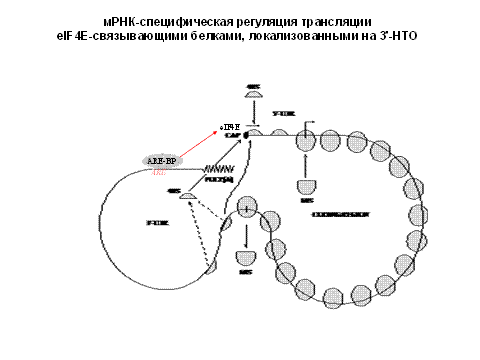
Взаимодействия AU-модулей (ARE)
3' нетранслируемой области:
Взаимодействия
ARE-BP – Cap
и ARE-BP – 5'-UTR
Самым главным является взаимудейтсвие АУ-богатых модулей. они очень распространены в 3"нетранслируемой области. Возможно, они взаимодействуют с 5"концом в циркулярном виде.
В основном снятие блока происходит из-за фосфорилирования белков.
Белок ЕВР. Он конкурирует за связывание с кэпом. блокируя посадку 40ки. его фосфорилирование - снимает этот блок.
|
|
|
Есть ВР, который сидит на определенном модуле в 3" нетранслируемой области.
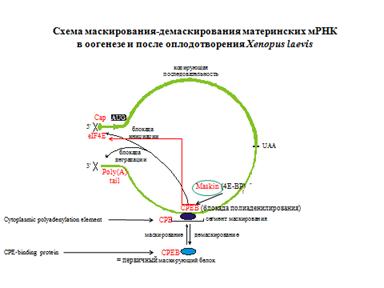
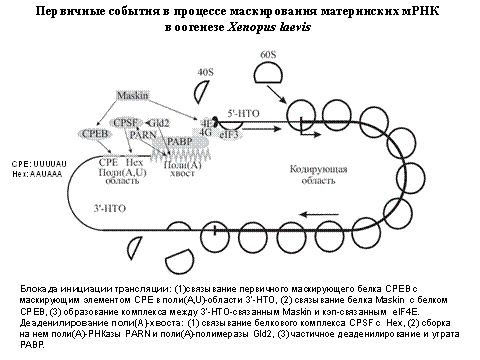
Это приводит к маскированию мРНК. Маскирование и демаскирование есть в оогенезе. Это происходит с помощью активаии киназ. Маскирование - недолгое явление. Это нужно для того, чтобы экзонуклеаза не ела мРНК.
МАСКИРОВАННАЯ мРНК
(1) Запасенная в процессе оогенеза мРНК ооцитов,
а также запасенная в сперматогенезе мРНК сперматоцитов.
(2) Маскирование-демаскирование мРНК в процессе эмбриогенеза и клеточной дифференцировки.
(3) Маскирование-демаскирование мРНК в ответ на физиологические и гормональные воздействия.
(4) Транспорт маскированной мРНК внутри клетки и
ее демаскирование в местах внутриклеточной локализации.
Особенности маскированных мРНК:
нетранслируемость;
стабильность, неатакуемость нуклеазами;
неподверженность полиаденилированию-деаденилированию
Оогенез:
Синтез и маскирование мРНК, кодирующих малую субъединицу рибонуклеотид-редуктазы, циклин А, etc., накопление мРНП (информосом) в цитоплазме.
Оплодотворение:
Демаскирование этих мРНК в цитоплазме,
переход из информосом в полисомы.
мРНП in vitro:
Демаскирование путем обработки солью (диссоциация белка мРНП).
Демаскирование с помощью антисмысловых РНК (“competitive unmasking assay”).
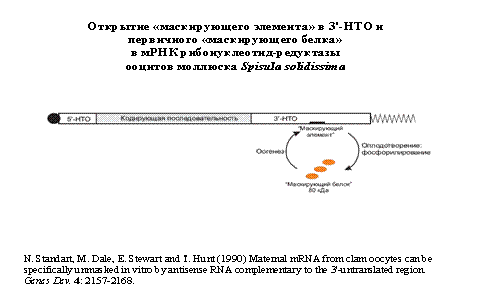
Открытие «маскирующего элемента» (“masking box”) в 3'-НТО:
N. Standart et al. (1990) Genes Dev. 4: 2157-2168.
Открытие маскирующего белка, специфически связывающегося с 3'-НТО:
N. Standart (1992) Semin. Dev. Biol. 3: 367-379.
Какая-то тетка выделяла маскированную мРНК из моллюска Spisula solidissima в атлантическом океане. она показала,что маскирование обеспечивается белками. Это было показано из того, что после удаления белков оказывалося, что мРНК рабочая.
Еще показали, что антисмысловая РНК к середке 3"нетранслируемой области репрессирует трансляцию. Это было очень неожиданно, потому что все искали маскирующий элементв 5" нетранслируемой области. Существует белок 80000кДа, который в оогенезе привлекается на маскирующий элемент. Фосфорилирование этого белка приводит к маскированию. За это присудили нобелевскую премию.
|
|
|
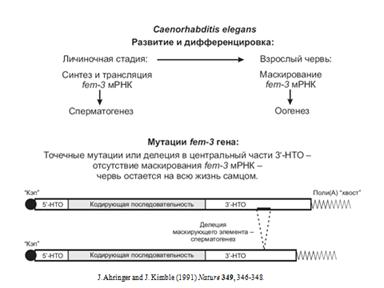
Есть нематода. Есть штаммы нематод с какой-то особренной fem-3 РНК. Они развиваются как самцы. Когда они вырастают, РНК маскируется и они превращаются в самок. при мутации в 3"последовательности, они на всю жизнь остаются самцами.
Дифференциировка эритробласта. Ретикулоцит плавает в крови. Происходит демаскирование РНК липоксигеназы. Она как-то приводит к тому, что происходит синтез гема - там не слышно как. при этом получается эритроцит.

Общая схема маскирования: есть нековалентная циклическая РНК. У нее есть сегмент маскирования. Там связывается маскирующий белок. Он каким-то образом влияет на блок инициации и блок деградации РНК.
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 1158; Нарушение авторских прав?; Мы поможем в написании вашей работы!