КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиологически активные соединения, взаимодействующие с серотониновым рецептором
|
|
|
|
Метаботропные глутаматные рецепторы
Антагонисты AMPA каинатных рецепторов
В настоящее время неизвестны специфические сильные антагонисты каинатных рецепторов. В тоже время известны либо сильные, но неселективные, либо селективные, но слабые антагонисты рецепторов AMPA. Например, диэтиловый эфир L‑глутаминовой кислоты является слабым, но селективным неконкурентным агонистом рецепторов AMPA.
Биоизостерическая замена терминальной карбоксильной группы L‑глутаминовой кислоты на динитрофенильную привела к высоко аффинным антагонистам рецепторов AMPA. Наличие свободных фенольных и аминогрупп важно для проявления активности, поскольку она резко уменьшается у кислород и азотзамещенных аналогов. Ситуация с селективными антагонистами для AMPA каинатных рецепторов резко отличается от таковой для рецептора NMDA, для которых изучено уже несколько поколений конкурентных антагонистов.
Семейство метаботропных глутаматных рецепторов было открыто совсем недавно. Активизация этих рецепторов приводит к запуску механизма с участием цепочки мембранных белков последовательно взаимодействующих друг с другом. На определенной стадии передача сигнала продолжается с помощью вторичных мессенджеров - молекул и ионов, которые в итоге вызывают изменения конфигурацию (конформацию) белков, участвующих в специальных клеточных процессах.
Семейство метаботропных глутаматных рецепторов подразделяются на три группы в зависимости от аналогий в строении их аминокислотной последовательности, механизмов передачи сигнала и чувствительности к действию различных агонистов.
Группа 1 принципиально отличается от групп 2 и 3. Взаимодействие агонистов с рецепторами группы 1 приводит к передаче возбуждения, а с группой 2 и 3 к блокированию возбуждения. Это лежит в основе стратегии поиска нейропротекторов, действующих на метаботропные глутаматные рецепторы.
|
|
|
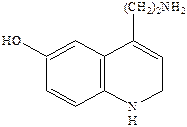
Серотонин или 5‑гидрокситриптамин (5НТ) является биогенным амином, образующимся в организме в результате превращения аминокислоты триптофана. Серотонин содержится в различных органах, тканях, тромбоцитах и в мозге. В нервной системе синтез серотонина происходит в цитоплазме нервных окончаний. Он накапливается в синаптических пузырьках, выделяется под влиянием нервных импульсов и взаимодействует со специфическими рецепторами, обозначаемыми как серотониновые (серотонин‑R‑ические). Серотониновые рецепторы содержатся в периферических тканях. Причем разделяются следующие подтипы: 5НТ1, 5НТ2, 5НТ3.
5НТ2 рецепторы содержатся в гладких мышцах стенок сосудов, в бронхах.
5НТ1 и 5НТ3 находятся в гладкой мускулатуре и в слизистой оболочке желудочно-кишечного тракта.
5НТ3 в центральной нервной системе и периферических тканях.
Известно, что физиологическая роль серотонина в головном мозге человека заключается в регуляции различных психоэмоциональных реакций. Кроме того, серотонин участвует в контроле терморегуляции, процессов сенсорного восприятия, в частности чувствительности к боли. В последующие годы было показано, что нарушение обмена серотонина и функций серотониновых регуляторов имеют отношение к патогенезу наркомании, алкоголизма и т.д. Эти факты и явились причиной возросшего интереса к изучению химических соединений, участвующих в нейропередаче серотонина.
Серотонин отличается чрезвычайно широким спектром биологической активности по отношению к центральной нервной системе и другим органам. С влиянием на биосинтез серотонина, его метаболизм и взаимодействие с рецепторами связан механизм действия ряда психотропных препаратов. Большая роль серотонину придается в механизме действия антидепрессантов.
|
|
|
Серотониновые рецепторы представляют собой белковые молекулы, встроенные в плазматические мембраны клеток и трансформирующие молекулярный сигнал серотонина и его аналогов в специальный ответ клетки. Действие большинства соединений используемых для лечения различных умственных расстройств, ожирения, мигрени основана на их участии в серотонин‑R‑ической передаче. Эти вещества выступают в роли агонистов или антагонистов соответствующих рецепторов. Очевидно, что антагонисты не обладают внутренней активностью. Возможно также существование частичных агонистов, не способных произвести максимально активных рецепторов, а следовательно, и максимальный биологический ответ независимо от применяемого количества.
Активация серотониновых рецепторов приводит к инициированию механизма с участием цепочки мембранных белков, последовательно взаимодействующих друг с другом. На определенной стадии передача сигнала продолжается с помощью вторичных мессенджеров («посыльные») – молекул или ионов, которые в итоге вызывают изменения конфигурации (конформации) белков, участвующих в специфических клеточных процессах.
Открытие различных подтипов серотониновых рецепторов привело к осознанию того факта, что большинство из известных лекарственных соединений, влиящих на серотонин‑R‑ическую передачу, действуют одновременно на несколько подтипов 5НТ рецепторов. Например, 5НТ3 рецептор образует ионный канал и передает сигнал непосредственно от нервных клеток за счет возникновения тока ионов, поэтому разработка селективных лигандов, приводящих к более эффективному лечению с меньшими побочными эффектами была и есть задача номер один нервных болезней, не говоря уже о том, что разработка селективных лигандов может привести к созданию принципиально новых форм терапии.
Серотониновые рецепторы найдены в высоких концентрациях в коре головного мозга, где, как полагают, они играют существенную роль в процессах возникновения эмоций. Лиганды, особенно агонисты, взаимодействующие с рецепторами были предложены в качестве терапевтических средств для лечения состояния страха, как обезболивающие средства и т.п.
|
|
|
Простейшие структурные модификации молекулы серотонина – введение различных заместителей в бензольное кольцо или ограничение конформационной подвижности в боковой цепи молекулы – привели к нескольким высоко аффинным агонистам 5НТ1 рецепторов, например,
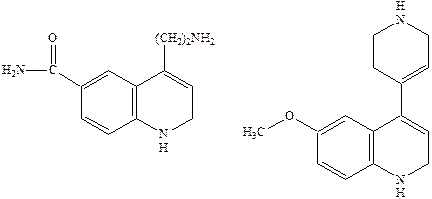
Однако указанные соединения как и сам серотонин не являются селективными и взаимодействуют с разными подтипами серотониновых рецепторов.
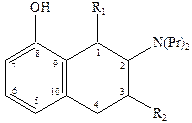
8‑гидрокси‑2‑дипропиламинотетралин оказался одним из наиболее активных селективных агонистов 5НТ1 рецепторов, причем полным агонистом является только (+)‑R‑энантиомер, который к тому же имеет приблизительно в двое большую активность.
Основные закономерности «структура-свойства» для тетралинов следующие:
1) Наличие гидроксильной группы в положении 8 необходимо для связывания. Обеспечивает 500‑кратную селективность к серотониновым рецепторам по сравнению с дофаминами.
2) Наличие второй гидроксильной группы в положении 5 или 7 приводит к резкому снижению селективности.
3) Возможна замена гидроксильной группы на метокси (-OCH3) и ацильную и некоторые другие, но не на карбоксильную, без потери аффинности и селективности, хотя при этом иногда сильно уменьшается агонистическая активность.
4) Оптимальным для аффинности, селективности и агонистической активности является наличие двух пропильных заместителей у атома азота, хотя некоторая вариация заместителей, например, Pr заменить на ‑CH2-CH=CHI, тоже дает хороший результат и приводит к значительному улучшению связывания полученного лиганда с рецептором.
5) Введение небольших заместителей R1 и R2 = ‑CH3 приводит к существенным различиям в активности полученных соединений.
6) Переход к три и тетрациклическим структурам, например, апоморфин, привел к созданию селективных сильных агонистов 5НТ1 рецепторов.
Основной проблемой в дизайне новых лигандов является обеспечение селективности, т.к. семейство серотониновых рецепторов характеризуется широким набором структурно похожих подклассов, и прогресс остается за рентгеноструктурными анализом, ибо только с его помощью можно уточнить компьютерные модели по докингу лидеров различных структурных классов.
|
|
|
|
|
Дата добавления: 2014-10-15; Просмотров: 629; Нарушение авторских прав?; Мы поможем в написании вашей работы!