КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Общее число клеток в отдельных слоях первичных, вторичных и третичных полей двигательной коры человека, млн
Высших млекопитающих
И премоторной областями на последовательных ступенях эволюции
(по данным Московского института мозга, 1958)
| Вид | Отношение | ||
| площади прецентральной области к общей поверхности коры | площади первичной двигательной коры (поля 4) к площади всей прецентральной области | площади премоторной области (поля 6) к площади всей прецентральной области | |
| Игрунка | 5,5 | ||
| Мартышка | 8,3 | ||
| Шимпанзе | 7,6 | 29,8 | 70,2 |
| Человек | 8,4 | 12,0 |
Таблица 13
{по данным Московского института мозга, 1960)
| Поле мозговой коры | Общее количество | Слои коры | ||
| Н-Ш | IV | V | ||
| — | 10,6 | |||
| — | 80,6 | |||
Анатомо-физиологический и критический анализы позволяют существенно уточнить характер участия премоторных зон в двигательных процессах. Как показали морфологические исследования, у человека в первичной двигательной зоне (4-е поле Бродмана) клетки в основном сконцентрированы в V (эфферентном) слое коры, в то время как количество клеток во II и III («ассоциативных») слоях коры сравнительно невелико, во вторичных, премоторных отделах двигательной коры (6-е поле Бродмана) число клеточных элементов в верхних слоях коры резко возрастает. В наиболее сложных — третичных областях коры (10-е поле Бродмана) — число клеточных элементов еще больше увеличивается (табл. 13).
Премоторная зона не имеет соматотопического строения, которым характеризуется проекционная зона двигательной коры, и ее раздражение, распространяясь на относительно большие территории (рис. 26), вызывает не изолированные сокращения мышечных групп, а комплексные движения, включающие повороты глаз и головы, а затем и туловища в противоположную сторону. Такие же адверзивные эпилептические припадки наблюдаются при наличии постоянных раздражающих факторов (например, рубцов) в премоторной области.
Итак, премоторные отделы коры являются аппаратом, приспособленным для интеграции эфферентных {двигательных) импульсов, и если постцентральные отделы коры обеспечивают пространственное распределение двигательных импульсов, то премоторные зоны коры, опираясь на нижележащие аппараты двигательных синергии, осуществляют превращение отдельных двигательных импульсов в последовательные кинетические мелодии, обеспечивая тем самым второй необходимый аспект организации сложных движений и двигательных навыков.
Сказанным выше определяются симптомы, возникающие при поражении премоторной зоны. Как это было показано еще в классических работах Фултона (1935, 1943), а затем и ряда других неврологов (Ферстер, 1936; Клейст, 1934; А.Р.Лурия, 1943, 1947, 1962, 1963, 1969), поражения премоторных отделов мозга не вызывают ни параличей, ни парезов противоположных конечностей. Их основным симптомом является отчетливое нарушение двигательных навыков {skilled movements). Клинически это проявляется в том, что у человека меняется почерк, каждый штрих буквы начинает требовать специального усилия, машинистка теряет быстроту и плавность своей работы, музыкант — плавность выполнения мелодий, квалифицированный рабочий оказывается не в состоянии автоматически выполнять серию операций, входящих в привычный двигательный акт.
Физиологически этот дефект выступает в том, что плавный процесс смены иннервации и денерваций нарушается; электромиографические импульсы, ведущие к возникновению движения, становятся патологически затянутыми (рис. 77).
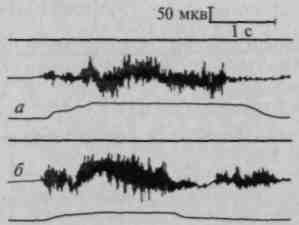
Рис. 77. Патологические изменения электромио- и механограммы при поражении премоторной области:
а — патология начала движения (трудности включения); б — патология конца движения (патологическая инертность) (по А.Я.Иошпа и Е.Д.Хомской)
В клинике такие явления хорошо известны под названием инертности двигательных стереотипов (рис. 78). Если предложить больному с поражением премоторной области отстучать сложный ритм, например ритм, включающий два сильных и три слабых удара, больной оказывается не в состоянии плавно переключаться с сильных ударов на слабые, его отстукивание приобретает отрывистый, дезавтоматизированный характер (А. Р. Лурия, 1943, 1963; Ф. М. Семерницкая, 1945; Б.Г.Спирин, 1966; и др.).
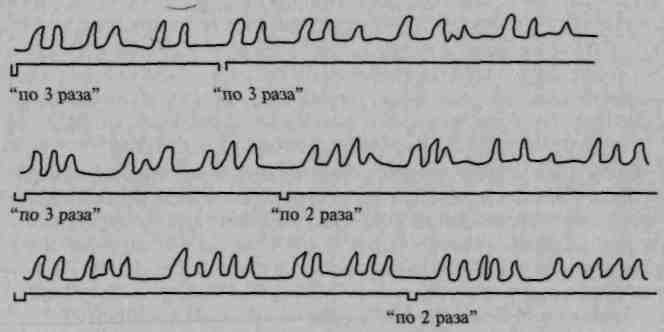
Рис. 78. Инертность элементарных двигательных стереотипов при
выполнении ритмических движений больным с поражением
премоторной области
В тех случаях, когда поражение премоторной области располагается глубоко, наблюдается иной симптом, имеющий большое принципиальное и диагностическое значение: тормозящая и модулирующая функции премоторной коры в отношении нижележащих подкорковых образований (подкорковых двигательных уз-
лов) устраняются, и начавшийся элемент движения не тормозится вовремя, бесконтрольно продолжаясь. Возникает явление, которое Н.А. Бернштейн (1947) назвал «зацикливанием» движения и которое широко известно под названием элементарной двигательной персеверации. На рисунках 79 и 80 приведены примеры таких нарушений движения.

Рис. 79. Двигательные персеверации у больного после пункции премоторной области, сопровождавшейся кровоизлиянием: а — рисунки и цифры; б — письмо

Рис. 80. Двигательные персеверации в послеоперационный период у больного с удалением опухоли (менигеомы) премоторной области:
а — рисование фигур (2—5-й дни после операции); б — рисование человечка (те же дни); в — рисование фигур и человечка на 7-й день после операции
Следует отметить, что наиболее отчетливо описываемые нарушения проявляются в контралатеральной очагу руке, но при поражении премоторной зоны левого (доминантного) полушария они часто проявляются в работе обеих рук. Отметим также, что во всех случаях у больных сохранны как намерение выполнить движение, так и общий план его выполнения, однако в своей исполнительной части движение высвобождается из-под влияния программы.
Остановимся еще на одном важном симптоме. В тех случаях, когда поражение располагается в нижних отделах премоторной зоны левого (доминантного) полушария, только что описанные нарушения плавного переключения с одного двигательного звена на другое (двигательные персеверации) начинают проявляться не только в движениях руки, но и в речи больного, приводя к своеобразному явлению, которое мы обозначили термином эфферентная (или кинетическая) моторная афазия (А. Р.Лурия, 1947, 1962, 1969, 1970).
В отличие от описанной выше афферентной (кинестетической) афазии нахождение нужных артикуляций и произношение изолированных речевых звуков не представляет для этих больных сколько-нибудь заметных трудностей. Отчетливые нарушения наступают тогда, когда больные переходят от одной артикуляции к другой (что имеет место при произношении любого слова или словосочетания). Процесс денервации предыдущей артикулемы и плавного переключения на последующую оказывается глубоко нарушенным, возникают явления патологической инертности раз возникшей артикуляции. Пытаясь произнести слово «муха», такой больной правильно артикулирует (губной) согласный, входящий в первый слог «му», но оказывается не в состоянии перейти на следующий (заднеязычно-гортанный) согласный в слоге «ха», и вместо требуемого слова произносит: «му...-м...м...му...ма».
Это нарушение кинетической организации артикуляций является основой той формы моторной афазии, которую в свое время описал Брока (1861) и которая по своим физиологическим механизмам резко отличается от описанной выше апрактической афферентной моторной афазии.
Следует отметить, что подобные нарушения выступают у больных с поражением нижних отделов левой премоторной зоны не только в устной речи, но и в письме как следствие нарушения плавности перехода от одного компонента слова к другому и патологической персеверации раз написанного слова (рис. 81).
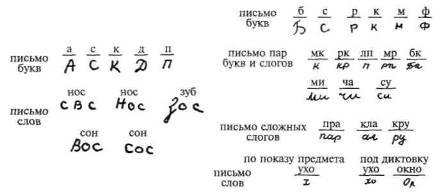
Рис. 81. Письмо больных с эфферентной (кинетической) моторной афазией
Таким образом, изучение функций премоторных отделов коры дает нам возможность вплотную подойти к механизмам одного из наиболее сложных явлений — нарушений моторной стороны речи.
Глава V
ЛОБНЫЕ ДОЛИ МОЗГА И РЕГУЛЯЦИЯ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ
Премоторные отделы мозговой коры входят в третий, основной, блок мозга, обеспечивающий программирование, регуляцию и контроль человеческой деятельности.
Как известно, лобные доли мозга, и в частности их третичные образования (к которым относится префронтальная кора), являются наиболее поздно сформировавшимся отделом больших полушарий. Едва намеченная у высших животных префронтальная кора заметно увеличивается у приматов, а у человека занимает до 25 % всей площади больших полушарий.
Префронтальные отделы мозга — или лобная гранулярная кора — состоят, главным образом, из клеток верхних (ассоциативных) слоев коры. Они имеют богатейшие связи как с верхними отделами ствола и образованиями зрительного бугра (рис. 45, а), так и со всеми остальными зонами коры (рис. 45, б). Таким образом, префронтальная кора надстраивается не только над вторичными отделами двигательной области, но фактически над всеми остальными образованиями большого мозга. Это обеспечивает двустороннюю связь префронтальной коры как с нижележащими структурами ретикулярной формации, модулирующими тонус коры, так и с теми образованиями второго блока мозга, которые обеспечивают получение, переработку и хранение экстероцептивной информации, что позволяет лобным долям регулировать общее состояние мозговой коры и протекание основных форм психической деятельности человека. Обе эти функции лобных долей мозга имеют настолько важное значение, что мы рассмотрим каждую из них по отдельности.
Функция лобных долей в организации поведения была подробно изучена многими авторами. Анализу их роли в организации поведения животных посвящены классические работы В.М.Бехтерева (1907), И.П.Павлова (1912-1913), П.К.Анохина (1949), Бианки (1895, 1921), Франца (1907), Джекобсона (1935), Малмо (1942), Прибрама (1954-1960), Росволда (1956 — 1959), Мишкина (1955—1958) и др. Анализ роли лобных долей в поведении человека был осуществлен в исследованиях Харлоу (1868), Вельт (1888), В.К.Хорошко (1912, 1921), Фейхтвангера (1923), Клейста (1943), Брикнера (1936), Райлендера (1939), Хэбба (1945), Халстеда (1947), Денни-Брауна (1951), А.Р.Лурия (1962, 1963, 1969), А.Р.Лурия, Е.Д.Хомской (1964, 1966), Е.Д.Хомской (1972) и др.
В следующей главе будут коротко изложены те данные, которые были получены в этих исследованиях.
1 ЛОБНЫЕ ДОЛИ И РЕГУЛЯЦИЯ СОСТОЯНИЙ АКТИВНОСТИ
Известно, что для протекания любых психических процессов необходим определенный тонус коры и что уровень этого тонуса зависит от поставленной задачи и от степени автоматизированной деятельности. Регуляция состояний активности составляет наиболее важную функцию лобных долей мозга.
Как уже говорилось ранее, состояние активного ожидания сигнала сопровождается появлением в лобных отделах мозга медленной биоэлектрической активности, которую Грей Уолтер назвал «волной ожидания» (рис. 48). Интеллектуальная деятельность также приводит к существенному увеличению в лобных отделах мозга количества синхронно работающих возбужденных пунктов (см. рис. 49). Эти факты свидетельствуют об особой роли лобных долей мозга в реализации сложных форм психической деятельности, осуществляемой при участии речевой системы.
Естественно ожидать, что при патологическом состоянии лобной коры должны нарушаться механизмы экстренной активации, опосредствованной речью.
Это положение было доказано многолетними работами Е. Д. Хомской и ее сотрудников (Е.Д.Хомская, 1960, 1961, 1966, 1972; Е.Ю.Артемьева, 1965; О.П.Барановская, 1968; А.Я.Иошпа, 1966; Э. Г. Симерницкая, 1966, 1970).
Остановимся на некоторых данных.
Известно, что каждый новый стимул вызывает ориентировочный рефлекс, выражающийся в соматических, сенсорных, вегетативных и других изменениях. Вегетативные компоненты ориентировочного рефлекса, проявляющегося в виде сужения периферических сосудов и расширения сосудов головы (Е.Н.Соколов, 1958, 1959; О.С.Виноградова, 1959), кожно-гальванической реакции и др., отмечаются до тех пор, пока раздражитель не становится привычным, и вновь восстанавливаются после введения задания, требующего различных операций с этим раздражителем (рис. 82).
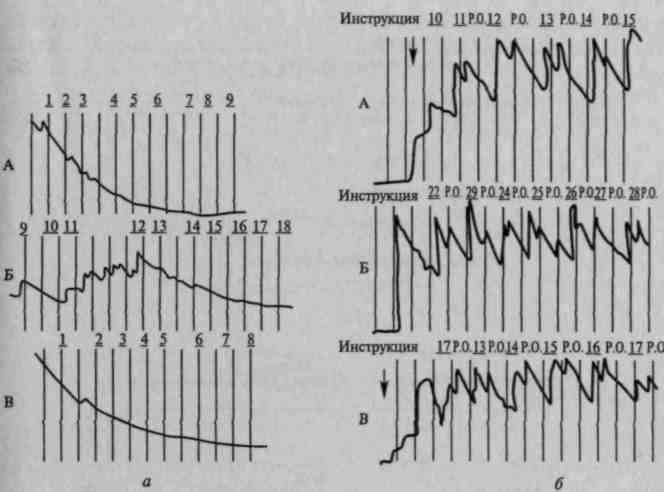
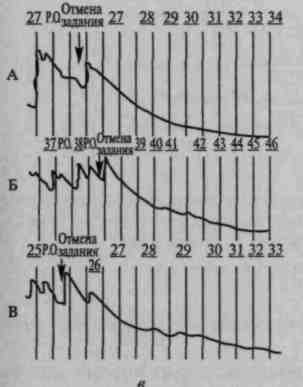
Рис. 82. Восстановление кожно-
гальванического компонента ориентировочного рефлекса при речевой инструкции, мобилизующей внимание у нормальных испытуемых (по Е.Д.Хомской):
а — угашение кожно-гальванической реакции на прерывистые звуки у трех испытуемых (А, Б, В); б — восстановление кожно-гальванических компонентов ориентировочной реакции после введения инструкции считать звуки; в — исчезновения этих реакций после отмены задания
При счете сигналов, оценке их по длительности и другим параметрам подобное восстановление вегетативных компонентов ориентировочного рефлекса, вызванного речевой инструкцией, отмечается у всех больных с поражением задних отделов мозга (рис. 83, а); однако у больных с поражением префронтальных отделов мозга (и особенно медиальных и базальных отделов лобной коры) оно либо становится очень нестойким, либо вообще не происходит (рис. 83, б). Это указывает на участие лобных долей мозга в регуляции процессов активации, лежащих в основе произвольного внимания.
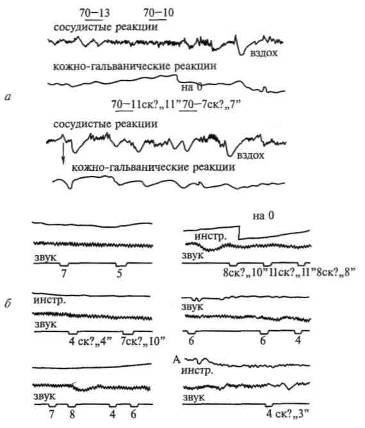
Рис. 83. Особенности регуляции вегетативных компонентов ориентировочного рефлекса с помощью речевой инструкции (считать звуки) у больных с поражением (по Е.Д.Хомской): а — задних; 6 — лобных отделов мозга
Аналогичные факты наблюдаются и при использовании электрофизиологических методов.
Известно, что в норме предъявление сколько-нибудь сложной задачи, требующей повышенного внимания, вызывает при соответствующих фоновых показателях отчетливые изменения электрической активности мозга, получившие названия реакции десинхронизации, или депрессии альфаритма: амплитуда электрических колебаний с частотой 8 — 13 Гц, известных под названием «альфа-ритма», снижается, в то время как амплитуда более частых колебаний возрастает.
Изменения частотного спектра электрической активности мозга при выполнении заданий, требующих повышения внимания (например, при задании считать количество сигналов, оценивать длительность сигналов и т.д.), обнаруживаются у большинства здоровых испытуемых и в той или иной мере у большинства больных с поражениями задних отделов коры (рис. 84, I, II). Однако у больных с поражениями лобных отделов мозга такие сдвиги, как показали наблюдения Е.Д.Хомской и О.П.Барановской (1966), Н.А. Филиппычевой и др., либо вообще не возникают, либо оказываются очень нестойкими или извращенными по знаку (рис. 84, III).
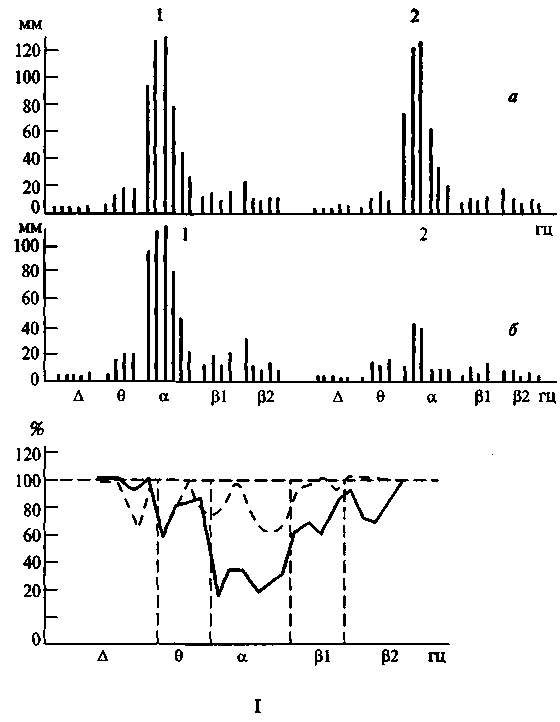
Рис. 84,1. Изменение частотного спектра электрической активности под влиянием речевой инструкции, вызывающей мобилизацию внимания в норме (по Е.Д.Хомской и О. П. Барановской).
В верхней части рисунка: а — спектр частот ЭЭГ в фоне (1) и во время действия индифферентных звуков (2); 6 — то же в фоне (1) и при выполнении инструкции считать звуки (2). В нижней части рисунка те же данные; изменение частотного спектра ЭЭГ по отношению к фону, принятому за 100%. Прерывистая линия — реакция на индифферентные звуки, сплошная линия — реакция на сигнальные звуки (выполнение инструкции считать звуки)
|
|
Дата добавления: 2014-11-25; Просмотров: 331; Нарушение авторских прав?; Мы поможем в написании вашей работы!