КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Мозжечок и процедурная память
|
|
|
|
МНОЖЕСТВЕННОСТЬ СИСТЕМ ПАМЯТИ
Современные исследования мозга, выполненные методом ПЭТ и функциональной МРТ, свидетельствуют, что актуализация следов памяти требует одновременной активации многих структур мозга, каждая из которых выполняет специфическую функцию по отношению к процессам памяти. Процессы памяти связывают с фронтальной, височной и париетальной корой, мозжечком, ба- зальными ганглиями, миндалиной, гиппокампом, неспецифической системой мозга.
Процесс формирования следа памяти характеризуется перемещением локусов активности по структурам мозга. Вновь формируемая энграмма в отличие от старого следа памяти представлена в мозге более широкой зоной активации.
Актуализация следа памяти предполагает обязательное появление активации в префронтальной коре, которая в режиме рабочей памяти обеспечивает считывание информации из основного
хранилища памяти — височной и теменной коры — и интеграцию ее на нейронах префронтальной коры. На пространственную локализацию активации влияет специфика информации, содержащейся в энграмме, которая выбирается в соответствии с решаемой задачей в системе целенаправленного поведения.
Высказана гипотеза о том, что след памяти через разное время после обучения реализуется разными по своему составу нейронными ансамблями (Греченко Т.Н., 1997). Как показало изучение динамики ассоциативного обучения у изолированных нейронов, более чем у 80% нейронов наблюдается отсроченное обучение. Эффект обучения проявляется через 5—40 мин после завершения процедуры обучения. Непосредственное или отсроченное обучение каждого нейрона привязано к определенному моменту времени и является устойчивой индивидуальной характеристикой нейрона при конкретном типе обучения. «Плавание» энграммы по структурам мозга (нейронным ансамблям) рассматривается как принцип организации памяти.
|
|
|
Мозжечок относится к многофункциональным структурам мозга. Среди его функций — сохранение равновесия, поддержание позы, регуляция и перераспределение мышечного тонуса, тонкая координация произвольных движений. В последние годы выявлена его способность одновременно с корой формировать все виды классических условных рефлексов. Благодаря связям клеток Пуркинье мозжечка со всеми сенсорными системами через мшистые, а затем через параллельные волокна, а также с нижней оливой, откуда поступают сигналы о всех совершаемых безусловных рефлексах, клетки Пуркинье представляют уникальную основу для конвергенции условного и безусловного сигналов.
Зачем в мозжечке параллельно неокортексу формируются условные рефлексы? Какую новую функцию по сравнению с корой берет на себя мозжечок? Предполагают, что мозжечок контролирует точность выполнения движений во времени, так как только мозжечок обладает способностью в любой момент времени заблокировать любую двигательную реакцию или, наоборот, дать ей возможность реализоваться. При поражении мозжечка клиницисты описывают явление дисметрии— плохое выполнение точных движений,
Причастность мозжечка к выработке классических условных рефлексов доказывается опытами с отключением холодом и разрушением его структур. Временное охлаждение моторных нейронов во время обучения блокирует выполнение условного и безус -
ловного рефлексов, но не нарушает сам процесс обучения. С восстановлением функций моторных нейронов можно обнаружить, что процедура обучения в этих условиях завершилась выработкой условного рефлекса, т.е. для процесса обучения исполнение условной и безусловной реакций необязательно. Если же во время выработки условного рефлекса холодом отключить кору и ядра мозжечка, обучение будет невозможным. Частичным разрушением ядер моста, откуда берут начало мшистые волокна, несущие афферентную информацию к клеткам Пуркинье, можно вызвать выпадение условных рефлексов на специфическую модальность. Замена условного сигнала электрической стимуляцией ядер моста приводит к более быстрому формированию условного рефлекса. Разрушение нижней оливы препятствует формированию условных рефлексов в мозжечке. Новые доказательства роли мозжечка в выработке классического условного рефлекса получены на мышах-мутантах, у которых через 2—4 недели после рождения происходит полная дегенерация коры мозжечка. Такие мыши способны лишь к частичному имплицитному обучению за счет ядерных структур мозжечка.
|
|
|
Значительный прогресс в изучении нейронных взаимодействий в мозжечке при выработке классических условных рефлексов был достигнут в работах Рихарда Томпсона и его коллег, доказавших, что условный мигательный рефлекс формируется не только в коре, но параллельно и в локальной зоне мозжечка. В качестве условного сигнала они использовали световой или звуковой раздражитель, а безусловным рефлексом служило мигание на обдувание воздухом роговицы кролика.
Основными клеточными элементами коры мозжечка являются клетки Пуркинье. Их дендриты восходят к поверхностным слоям, а аксоны идут к нейронам ядер мозжечка (рис. 26). Кроме клеток Пуркинье, в коре мозжечка имеются зернистые (или клетки-зерна) и корзинчатые клетки. Афферентная информация поступает в мозжечок к зернистым клеткам от мшистых волокон (аксонов нейронов моста). Клетки-зерна посылают параллельные волокна к ден-дритам каждой клетки Пуркинье, образуя на них множество синапсов. Параллельно клетки-зерна образуют синапсы и на корзин- чатых клетках (интернейронах), оканчивающихся тормозными синапсами на соме клетки Пуркинье. Активация клеток Пуркинье создает торможение у нейронов ядер мозжечка посредством ГАМК-тормозного медиатора и как следствие — торможение нейронов красного ядра, управляющих двигательными рефлексами. На дендриты клеток Пуркинье конвергируют не только параллельные волокна зернистых клеток, но и лазящие (или ползующие} волок-
|
|
|
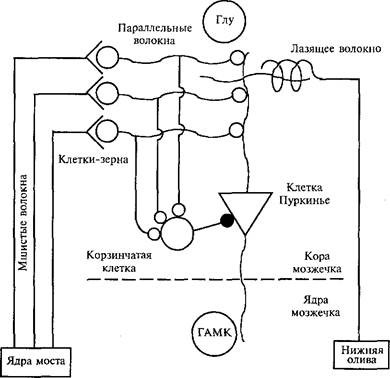
Рис. 26. Схема нейронной сети в мозжечке, обеспечивающей функционирование условного мигательного рефлекса.
на — аксоны нейронов нижней оливы. Имеется около 15 млн клеток Пуркинье. Каждая из них имеет возбуждающие синаптические контакты только с одним лазящим волокном и много тысяч синапсов — от параллельных волокон.
Таким образом, на дендритах каждой клетки Пуркинье сходятся два потока влияний: от самых разных афферентов (параллельные волокна) и от нижней оливы через одно лазящее волокно.
Сигнал, формирующийся в нижней оливе от безусловного рефлекса, т.е. сигнал от подкрепления, передается лазящему волокну, которое осуществляет селективный выбор определенной клетки Пуркинье, действуя по принципу: одно лазящее волокно — одна клетка Пуркинье. Условный раздражитель представлен на клетках Пуркинье возбужденным параллельным волокном. Подобная конвергенция волокон имеет место и на нейронах ядер мозжечка.
| П •ж мс | /ть ус. нтеля эзжечк | лобного раздра-к структурам а | Путь безусловного раздражителя к структурам мозжечка | |||||
| Кора мозжечка (клетки Пуркинье) | ||||||||
| Мшис волок | тые сна | Лазящие волокна | ||||||
| 1 ' | ||||||||
| Ядр мозже | а | |||||||
| я Ьй и -§• й> & е 0 2 и >. та 1-1 >, ^ | чка | |||||||
| Красное ядро | Нижняя олива | |||||||
| Ядра моста | ||||||||
| ' | ||||||||
| . | . | |||||||
| Мотонейроны | Ядра лицевых | |||||||
| та и ^ и ^ и ь 0 0 х ЕС 0 >. м <и ю ' ' я 1- >1 ^ | МЫШЦ * | |||||||
| г | Ретикулярная формация | |||||||
| /» | Акустическое ядро | |||||||
| Яд троит не | ра | |||||||
| < | Мышца третьего века | |||||||
| ичного | ||||||||
| рва | ||||||||
| р | Звук (условный аздражитель) | Воздух (безусловный раздражитель) | ||||||
| Реакция |
Рис. 27. Схема взаимодействия мозжечка и структур мозга, причастных к выработке условного мигательного рефлекса.
|
|
|
Обдувание воздухом роговицы возбуждает ядра тройничного нерва и VI и VII пары ядер лицевых мышц. Сигнал достигает мотонейронов, управляющих движением мышц века, и вызывает мигательную реакцию. От ядер тройничного нерва к ядрам лицевых мышц идет дополнительный путь через РФ, а также путь к мозжечку через нижнюю оливу (рис. 27). Нейроны мозжечка имеют только один выход — пучок аксонов клеток Пуркинье.
Установлено, что клетки Пуркинье характеризуются тонической спонтанной активностью. Ее увеличение означает усиление их тормозных влияний на нейроны ядер мозжечка и красное ядро. Оно уменьшается со снижением активности клеток Пуркинье. Тем самым создаются условия для активации моторных центров и появления двигательной реакции.
При выработке условного рефлекса в мозжечке возникает избирательное торможение определенных клеток, Пуркинье по принципу «торможение торможения», в результате чего из-под тормозного контроля высвобожается определенный безусловный рефлекс.
Плохое выполнение точных движений у пациентов с дисфункцией мозжечка сочетается с дефектом в когнитивной сфере. У них нарушены последовательность и согласованное исполнение когнитивных операций, из-за этого страдают генерация идей, формулирование гипотез. Таким образом, можно говорить не только о двигательной, но и о когнитивной дисметрии, возникающей в результате нарушений функций мозжечка. Мозжечок работает в единой системе с фронтальной корой и таламусом. Префронтальная кора, мозжечок и таламус, по данным ПЭТ, активируются одновременно. Это объясняют тем, что функция префронтальной коры, которая задает программу действий, дополняется функцией мозжечка, который контролирует точное ее исполнение во времени. Возможно, что дезорганизация мышления у шизофреников связана не только с нарушением рабочей памяти, но и с дисфункцией циркуляции процессов в системе Префронтальная кора—таламус— мозжечок. По данным, полученным с помощью ПЭТ, у таких пациентов уровень кровотока в этих структурах по сравнению с нормой снижен.
|
|
|
|
|
Дата добавления: 2014-12-10; Просмотров: 1021; Нарушение авторских прав?; Мы поможем в написании вашей работы!