КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение синапса. Синаптическая передача возбуждения ….….. 23 2 страница
|
|
|
|
Таким образом, выходящие из клетки катионы калия совместно с избытком катионов натрия во внеклеточной жидкости создают положительный потенциал на наружной поверхности мембраны покоящейся клетки.
В состоянии покоя плазматическая мембрана нервных клеток мало проницаема, а мембрана мышечных клеток хорошо проницаема для анионов хлора. Анионы хлора, которых больше во внеклеточной жидкости, диффундируют внутрь мышечной клетки и несут с собой отрицательный заряд. Полного уравнивания концентраций ионов хлора снаружи и внутри клетки не происходит, т.к. этому препятствует сила электрического взаимного отталкивания одноименных зарядов. Создаётся хлорный равновесный потенциал, при котором вход ионов хлора в клетку и их выход из неё находятся в равновесии.
Мембрана клетки практически непроницаема для крупных анионов органических кислот и молекул белков. Поэтому они остаются в цитоплазме и обеспечивают отрицательный потенциал на внутренней поверхности мембраны покоящейся нервной клетки. В мышечных клетках в создание отрицательного потенциала большой вклад вносят поступающие внутрь клетки анионы хлора.
Важнейшее значение мембранного потенциала покоя состоит в том, что он создаёт электрическое поле, которое воздействует на макромолекулы мембраны и придаёт их заряженным группам определённое положение в пространстве. Особенно важно то, что это электрическое поле обусловливает закрытое состояние активационных «ворот» натриевых каналов и открытое состояние их инактивационных «ворот» (рис. 10, А). Этим обеспечивается состояние покоя клетки и готовности её к возбуждению. Даже относительно небольшое уменьшение мембранного потенциала покоя открывает активационные «ворота» натриевых каналов, что выводит клетку из состояния покоя и даёт начало возбуждению.
|
|
|
Потенциал действия
При действии на клетку различных раздражителей мембранный потенциал покоя начинает уменьшаться, т.е. происходит деполяризация мембраны клетки. С увеличением силы раздражения деполяризация мембраны нарастает. Однако если сила раздражения не достигла определённой (пороговой) величины, то прекращение раздражения приводит к быстрому восстановлению потенциала покоя. В нервных и мышечных клетках при слабом (подпороговом) раздражении, не вызывающем ответной реакции, уменьшение потенциала покоя ограничено небольшим участком мембраны в том месте, где наносится раздражение. Такое небольшое уменьшение потенциала мембраны получило название местного потенциала, или локального ответа.
При достижении достаточно большой (пороговой) силы раздражения возникает быстрое кратковременное изменение величины и полярности заряда мембраны клетки, которое получило название потенциала действия, или волны возбуждения (рис. 11), или нервного импульса (для нервных клеток). Потенциал действия возникает при деполяризации мембраны нервной и поперечно-полосатой мышечной клетки, как правило, примерно до –50 мВ.
Причиной возникновения местного потенциала, а затем и потенциала действия является раскрытие активационных «ворот» натриевых каналов (рис. 10, Б) и поступление ионов натрия внутрь клетки. При нарастании силы раздражения до пороговой величины увеличение количества открытых каналов идёт медленно и происходит формирование местного потенциала.
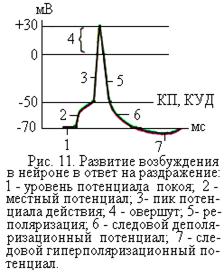 Когда заряд на внутренней поверхности мембраны клетки достигнет примерно –50 мВ, лавинообразно возрастает раскрытие натриевых каналов мембраны. Этот заряд на мембране клетки, при котором происходит раскрытие практически всех натриевых каналов, получил название критического потенциала. Состояние мембраны в момент лавинообразного раскрытия натриевых каналов называют критическим уровнем деполяризации мембраны (рис. 11). Разность между величинами критического потенциала и потенциала покоя называется пороговым потенциалом. Пороговый потенциал показывает, на какую величину надо уменьшить мембранный потенциал покоя, чтобы возник потенциал действия. При ухудшении функционального состояния клетки пороговый потенциал возрастает.
Когда заряд на внутренней поверхности мембраны клетки достигнет примерно –50 мВ, лавинообразно возрастает раскрытие натриевых каналов мембраны. Этот заряд на мембране клетки, при котором происходит раскрытие практически всех натриевых каналов, получил название критического потенциала. Состояние мембраны в момент лавинообразного раскрытия натриевых каналов называют критическим уровнем деполяризации мембраны (рис. 11). Разность между величинами критического потенциала и потенциала покоя называется пороговым потенциалом. Пороговый потенциал показывает, на какую величину надо уменьшить мембранный потенциал покоя, чтобы возник потенциал действия. При ухудшении функционального состояния клетки пороговый потенциал возрастает.
|
|
|
Через открытые каналы ионы натрия поступают из внеклеточной жидкости, где концентрация их больше, внутрь клетки, где их меньше, и несут с собой положительный заряд. Это приводит не только к быстрой деполяризации, т.е. к уменьшению до нуля отрицательного заряда на внутренней поверхности мембраны клетки, но и к возникновению там положительного заряда. Быстрые изменения величины и полярности заряда мембраны получили название пика потенциала действия, или просто потенциала действия (рис. 11). Фаза потенциала действия, во время которой внутренняя поверхность мембраны клетки имеет положительный заряд, называется овершут (инверсия).
Однако поступление ионов натрия в клетку быстро прекращается, т.к. продолжающаяся и увеличившаяся деполяризация мембраны приводит к закрытию инактивационных «ворот» (рис. 10, В). Таким образом, поступление ионов натрия в клетку длится тот короткий промежуток времени, пока открыты активационные и не закрылись инактивационные «ворота». Если бы в натриевых каналах при деполяризации активационные и инактивационные «ворота» срабатывали одновременно, то ионы натрия не смогли бы поступить в клетку. Однако в связи с тем, что скорость срабатывания активационных «ворот» выше, то активация обгоняет инактивацию, и натриевые каналы кратковременно находятся в открытом состоянии.
Закрытие (инактивация) натриевых каналов – это очень важный механизм, который способствует прекращению пика потенциала действия и развитию временной невозбудимости клетки, или рефрактерности. Инактивация натриевых каналов устраняется после реполяризации мембраны.
После закрытия натриевых каналов резко увеличивается выход ионов калия из клетки через калиевые каналы. Катионы калия устремляются по градиенту концентрации из клетки с большой скоростью, т.к. на них, кроме выталкивающей осмотической силы, в течение короткого промежутка времени действует притягивающая сила отрицательного электрического заряда на наружной поверхности мембраны. В результате выхода из клетки положительных ионов калия отрицательный заряд на наружной поверхности мембраны уменьшается до нуля (реверсия). Затем наружная поверхность мембраны вновь приобретает положительный заряд, а внутренняя поверхность мембраны – отрицательный заряд, т.е. происходит процесс реполяризации, или восстановления первоначальной полярности и величины мембранного потенциала.
|
|
|
Продолжительность потенциала действия в нерве составляет примерно 1 мс, в поперечнополосатой мышечной клетке – 10 мс.
Следовые потенциалы
Восстановление мембранного потенциала покоя после осуществления пика потенциала действия происходит не сразу, а после следовых процессов, выражающихся в характерных изменениях мембранного потенциала. Эти изменения, следующие за пиком потенциала действия, называют следовыми потенциалами. Различают два вида следовых потенциалов – следовой деполяризационный и следовой гиперполяризационный потенциал (рис. 11). Величина следовых потенциалов обычно не превышает нескольких милливольт, а длительность их у различных нервных волокон составляет от нескольких миллисекунд до нескольких сотен миллисекунд.
Следовой деполяризационный потенциал наблюдается в тот период, когда реполяризация, обусловленная выходом ионов К+ из клетки, начинает происходить медленнее. Это вызвано тем, что по мере выхода катионов калия из клетки, ослабевает осмотическая сила выталкивания и усиливается электрическая сила отталкивания катионов от положительно заряженной наружной поверхности мембраны. Во время следового деполяризационного потенциала мембрана клетки имеет меньшую величину заряда, чем в покое, т.е. она слегка деполяризована. В тот период, когда величина деполяризационного потенциала находится между величиной потенциала покоя и величиной критического потенциала, клетка обладает повышенной возбудимостью. В это время она может ответить потенциалом действия и ответной реакцией даже на более слабые раздражители, на которые в обычных условиях она не отвечает.
|
|
|
В безмякотных нервных волокнах вслед за пиком потенциала действия развивается следовой гиперполяризационный потенциал. Он обусловлен более длительным сохранением повышенной проницаемости мембраны для ионов калия. Вследствие этого катионов калия выходит из клетки больше и наружная поверхность мембраны приобретает на какое-то время более положительный заряд, а внутренняя поверхность мембраны – более отрицательный заряд, чем в покое. Наблюдается период гиперполяризации мембраны, во время которого клетка менее возбудима, чем в покое.
В мякотных нервных волокнах следовые потенциалы имеют более сложный характер. Следовой деполяризационный потенциал может переходить в следовой гиперполяризационный потенциал и лишь после этого происходит полное восстановление потенциала покоя. Следовые потенциалы в значительно большей мере, чем пики потенциалов действия, чувствительны к изменениям ионного состава среды, кислородному снабжению клетки и т.д. Характерной особенностью следовых потенциалов является их способность изменяться в процессе ритмического раздражения.
На осуществление одного потенциала действия вместе со следовыми потенциалами расходуется очень мало ионов. Разницы концентрации ионов калия и натрия внутри и снаружи нервного волокна может хватить на 500 000 импульсов. Но для длительной активности нейронов необходимо восстанавливать градиенты ионных концентраций, что обеспечивается работой натрий-калиевого насоса мембраны клетки.
ПРОВЕДЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ
Распространение нервного импульса по безмякотным нервным
волокнам
Распространение потенциалов действия по ткани, особенно нервного импульса по нервам, является самым быстрым и точно адресованным способом передачи информации в организме. Скорость передачи нервного импульса в быстропроводящих волокнах двигательных нервов (тип волокна Аα) достигает 120 м/с. Другие способы передачи информации гораздо медлительнее: гуморальный не превышает 0,5 м/с (скорость тока крови в аорте), аксонный транспорт веществ от тела нейрона к окончаниям аксона не превышает 40 см в сутки.
Распространение потенциалов действия, или проведение возбуждения, осуществляется по мембране нервных и мышечных клеток. Проведение нервных импульсов по безмякотным и мякотным нервным волокнам имеет свои особенности.
При нанесении раздражения на безмякотное нервное волокно, не содержащее в своей оболочке электроизолирующего вещества миелина, в раздражаемом участке изменяется ионная проницаемость мембраны. Раскрываются активационные «ворота» натриевых каналов и ионы натрия поступают внутрь нервного волокна. Развивается потенциал действия и мембрана возбуждённого участка заряжается снаружи отрицательно, а внутри положительно (рис. 12).
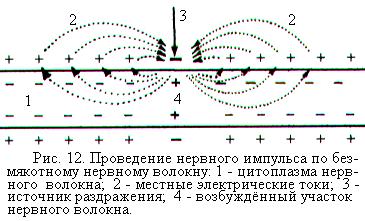 Входящий внутрь волокна положительный ток, создаваемый катионами натрия, должен выходить через мембрану наружу, чтобы замкнуть электрическую цепь. Соседние невозбуждённые участки мембраны нервного волокна имеют на наружной поверхности положительный заряд, а на внутренней – отрицательный. Между возбуждённым и невозбуждёнными участками мембраны возникают местные электрические токи, которые, проходя через невозбуждённые участки мембраны, раздражают их. В результате в них изменяется ионная проницаемость мембраны, эти участки возбуждаются и в них возникает потенциал действия. Процесс повторяется и таким образом возбуждение проводится в обе стороны по нервному волокну от первоначального места нанесения раздражения.
Входящий внутрь волокна положительный ток, создаваемый катионами натрия, должен выходить через мембрану наружу, чтобы замкнуть электрическую цепь. Соседние невозбуждённые участки мембраны нервного волокна имеют на наружной поверхности положительный заряд, а на внутренней – отрицательный. Между возбуждённым и невозбуждёнными участками мембраны возникают местные электрические токи, которые, проходя через невозбуждённые участки мембраны, раздражают их. В результате в них изменяется ионная проницаемость мембраны, эти участки возбуждаются и в них возникает потенциал действия. Процесс повторяется и таким образом возбуждение проводится в обе стороны по нервному волокну от первоначального места нанесения раздражения.
Местные токи выходят из волокна наружу в области наименьшего сопротивления мембраны. Расстояние, на которое распространяется каждый «виток» местных токов по волокну, тем больше, чем ниже сопротивление цитоплазмы и выше сопротивление мембраны. В тонких безмякотных нервных волокнах это расстояние, т.е. протяжённость возбуждённого участка, тем больше, чем толще волокно, но не превышает 1 мм.
Чем дальше от точки раздражения, тем слабее местные токи. Поэтому на расстоянии больше 1 мм они не могут проходить через мембрану и вызывать образование новых потенциалов действия. Если источник раздражения убрать, то в первоначальном месте нанесения раздражения благодаря действию натрий-калиевого насоса восстановится мембранный потенциал покоя.
В безмякотных нервных волокнах местные токи проходят последовательно практически через каждый соседний участок мембраны волокна, т.к. длина их «витка» не превышает 1 мм. Поэтому скорость распространения нервного импуьса невелика (0,5 - 3 м/с) и его передача требует бóльшего расхода энергии, чем при распространении импульсов по мякотным нервным волокнам.
Распространение нервного импульса по мякотным нервным
волокнам. Типы нервных волокон
В мякотных нервных волокнах, покрытых электроизолирующей миелиновой оболочкой, местные токи могут проходить через мембрану только в перехватах Ранвье – участках волокна, лишённых миелина. Мембрана перехватов Ранвье специализирована для возникновения потенциалов действия. Здесь плотность натриевых каналов в 100 раз выше, чем в безмякотных нервных волокнах, и достигает 12 000 на 1 мкм2. Поэтому в мякотных нервных волокнах распространение нервного импульса осуществляется скачкообразно: возбуждение как бы перескакивает от одного перехвата Ранвье к другому (рис. 13). Расстояние между перехватами во много раз больше (1000 – 2000 мкм), чем длина самого перехвата Ранвье (1 мкм). Поэтому на передачу импульсов по мякотным нервным волокнам расходуется меньше энергии, чем в безмякотных, а скорость распространения импульсов гораздо выше.
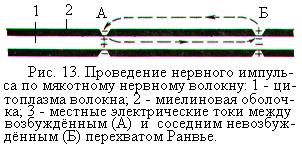 Величина потенциала действия, возникшего в одном перехвате Ранвье, в 5-10 раз больше той пороговой величины, которая необходима для возбуждения соседнего перехвата. Поэтому потенциал действия, возникший в одном перехвате, способен вызвать возбуждение не только в находящемся рядом, но и в соседних 2-3 перехватах. Это создаёт гарантию проведения импульсов по нервному волокну, даже если 1-2 ближайших перехвата Ранвье повреждены.
Величина потенциала действия, возникшего в одном перехвате Ранвье, в 5-10 раз больше той пороговой величины, которая необходима для возбуждения соседнего перехвата. Поэтому потенциал действия, возникший в одном перехвате, способен вызвать возбуждение не только в находящемся рядом, но и в соседних 2-3 перехватах. Это создаёт гарантию проведения импульсов по нервному волокну, даже если 1-2 ближайших перехвата Ранвье повреждены.
Скорость распространения импульсов зависит от диаметра нервных волокон, наличия и толщины миелиновой оболочки. Нервы у человека состоят из трёх основных типов волокон: А, В и С. Тип А включает наиболее толстые хорошо миелинизированные афферентные и эфферентные нервные волокна и делится на 4 группы в зависимости от диаметра волокна и скорости проведения возбуждения. Волокна группы Аα имеют диаметр 13-22 мкм и скорость проведения импульсов 70-120 м/с; группы Аβ – диаметр 8-13 мкм и скорость 40-70 м/с; группы Аγ – диаметр 4-8 мкм, скорость 15-40 м/c; группы Аδ – диаметр 1-4 мкм и скорость проведения возбуждения 5-15 м/с.
Тип В включает в себя слабомиелинизированные волокна автономной нервной системы, имеющие диаметр 1-3 мкм и скорость проведения возбуждения 3-14 м/с. В тип С объединены безмякотные волокна автономной нервной системы диаметром 0,5-1 мкм и со скоростью проведения импульсов 0,5-2 м/с.
Строение синапса. Синаптическая передача возбуждения
По нервному волокну одной нервной клетки импульсы могут распространяться в обе стороны от того места, где на это волокно действует раздражитель. Однако передача импульсов с нейрона на нейрон или с нейрона на клетки рабочего органа может осуществляться только в одном направлении. Односторонность передачи возбуждения обеспечивается особыми образованиями, которые английский физиолог Ч. Шеррингтон назвал синапсами.
Синапс – это место контакта между двумя нейронами, или между нейроном и клеткой рабочего органа, специально предназначенное для передачи нервных импульсов. Синапсы классифицируют: 1) по месторасположению; 2) по оказываемому действию; 3) по способу передачи сигналов.
По месторасположению синапсы бывают: а) аксо-соматические, т.е. образованные аксоном одного нейрона на теле, или соме, другого нейрона; б) аксо-дедритические, т.е. образованные аксоном одного нейрона на дендрите другого нейрона; в) аксо-аксональные, образованные аксоном одного нейрона на аксоне другого нейрона; г) дендро-дендритические, образованные дендритом одного нейрона на дендрите другого нейрона. Синапсы между окончаниями аксона и иннервируемыми мышечными клетками получили название нервно-мышечных синапсов, или концевых пластинок.
По оказываемому действию синапсы бывают возбуждающие и тормозные. По способу передачи сигналов различают электрические синапсы, в которых сигналы передаются электрическим путём, и химические синапсы, в которых передатчиками сигнала, или медиаторами, являются особые биологически активные химические вещества. Существуют также смешанные электрохимические синапсы. Почти все синапсы в организме человека являются химическими. На теле и отростках одного нейрона может быть несколько тысяч синапсов.
Во всех синапсах имеются такие составные компоненты, как пресинаптический отдел, постсинаптический отдел и разделяющая их синаптическая щель (рис 14).
Пресинаптический отдел химического синапса образован расширенным окончанием нервного волокна, которое перед синапсом лишено миелиновой оболочки. В пресинаптическом отделе синтезируется медиатор, который накапливается в синаптических пузырьках. Со стороны синаптической щели пресинаптический отдел покрыт пресинаптической мембраной. В ней имеются электровозбудимые кальциевые каналы, которые открываются под воздействием приходящего нервного импульса, в результате чего ионы кальция поступают в пресинаптический отдел из внеклеточной жидкости.
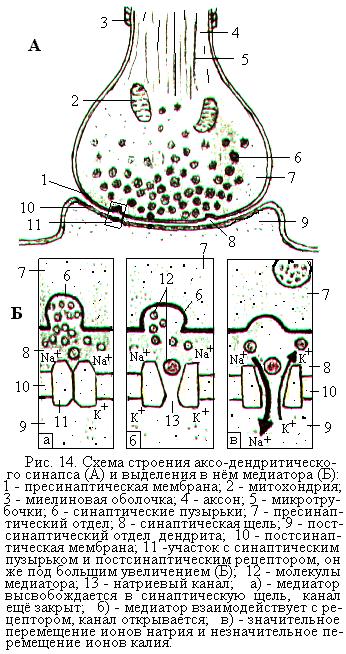 Поступление ионов кальция, вызванное нервным импульсом, приводит к перемещению синаптических пузырьков с медиатором к пресинаптической мембране и практически одновременному выделению медиатора из 100-200 пузырьков в синаптическую щель. Каждый из пузырьков содержит одну порцию, или квант, медиатора. Квант состоит из нескольких тысяч молекул медиатора, которые выходят в синаптическую щель за 1-2 мс. Для выхода в щель каждого кванта медиатора нужны 4 иона Са2+.
Поступление ионов кальция, вызванное нервным импульсом, приводит к перемещению синаптических пузырьков с медиатором к пресинаптической мембране и практически одновременному выделению медиатора из 100-200 пузырьков в синаптическую щель. Каждый из пузырьков содержит одну порцию, или квант, медиатора. Квант состоит из нескольких тысяч молекул медиатора, которые выходят в синаптическую щель за 1-2 мс. Для выхода в щель каждого кванта медиатора нужны 4 иона Са2+.
Размеры синаптической щели очень малы, примерно 20-50 нм, и молекулы медиатора путём диффузии быстро, менее чем за 1 мс, достигают постсинаптической мембраны. Эта мембрана покрывает со стороны синаптической щели постсинаптический отдел и имеет специфические утолщения – постсинаптические рецепторы, обладающие повышенной чувствительностью к медиатору (рис. 14, Б). Молекулы медиатора воздействуют на мембрану постсинаптических рецепторов и вызывают в ней открытие каналов, проницаемых для ионов натрия и калия. В результате происходит частичная деполяризация постсинаптической мембраны, которая получила название возбуждающего постсинаптического потенциала (ВПСП).
ВПСП имеет разную величину для разных синапсов, но чаще всего он равен 10-20 мВ. ВПСП соседних синапсов на теле одного нейрона суммируются между собой. Так же суммируются ВПСП, быстро возникающие друг за другом в одном синапсе. Когда в результате такой суммации общая деполяризация постсинаптической мембраны достигнет примерно 60 мВ, в постсинаптическом отделе возникает потенциал действия (нервный импульс), который распространяется по второй клетке электрическим путём. Не вступившая во взаимодействие с постсинаптическим рецептором часть медиатора разрушается специальным ферментом или поступает обратно через пресинаптическую мембрану в пресинаптический отдел.
Таким образом, через химический синапс возбуждение передаётся химическим путём с помощью медиатора только в одном направлении – от пресинаптического отдела к постсинаптическому. Скорость передачи возбуждения через синапс примерно в 200 раз меньше, чем по нервному волокну. Это получило название синаптической задержки. Синаптическая задержка составляет около 0,2 – 0,5 мс, причём основная часть этого времени тратится на процесс выделения медиатора в синаптическую щель.
В состоянии относительного покоя, даже когда в пресинаптический отдел не поступают нервные импульсы, тем не менее, происходит самопроизвольный выброс одиночных квантов медиатора в синаптическую щель. Это вызывает кратковременную слабую деполяризацию постсинаптической мембраны, которая получила название миниатюрного возбуждающего постсинаптического потенциала (МВПСП). Одиночный квант медиатора приводит к открытию примерно на 1 мс одного ионного канала в постсинаптическом рецепторе. При этом возникает МВПСП величиной 0,1 – 0,2 мВ.
Синапсы, в которых поступивший нервный импульс вызывает возбуждение второй клетки, называются возбуждающими. Медиаторами в возбуждающих синапсах чаще всего являются такие химические вещества как ацетилхолин, норадреналин и др.
Синапсы, в которых поступивший нервный импульс вызывает торможение второй клетки, называют тормозными. В них выделяются тормозные медиаторы – гаммааминомасляная кислота (ГАМК), глицин и др. Тормозной медиатор, взаимодействуя с постсинаптическим рецептором, открывает в нём избирательные каналы для ионов хлора. В результате ионы хлора поступают из внеклеточной среды, где их больше, в цитоплазму клетки, где их меньше, и несут с собой отрицательный заряд. Происходит не уменьшение, а увеличение заряда на мембране (гиперполяризация), что приводит к торможению клетки, на которой находится тормозной синапс. Величина, на которую увеличивается заряд постсинаптической мембраны в результате действия тормозного медиатора, называется тормозным постсинаптическим потенциалом (ТПСП).
В зависимости от того, какой по химической природе медиатор выделяется в синаптическую щель, синапсы подразделяют на холинергические (медиатор – ацетилхолин), норадренергические (медиатор – норадреналин), ГАМК-ергические (медиатор – ГАМК), серотонинергические (медиатор – серотонин) и т.д.
Рефлекторная деятельность нервной системы
Основной формой деятельности нервной системы является осуществление рефлексов. Рефлексы – это реакции организма, которые возникают в ответ на раздражение рецепторов и осуществляются при обязательном участии нервной системы. Благодаря рефлекторным реакциям происходит постоянное взаимодействие организма с окружающей средой, объединение и регуляция деятельности всех его органов и тканей.
 Путь, по которому проходит нервный импульс при осуществлении рефлекса, называют рефлекторной дугой. В самые простые рефлекторные дуги входят только по два нейрона, в более сложные – по три, а в большинстве рефлекторных дуг насчитывается ещё больше нейронов. Примером двухнейронной рефлекторной дуги является дуга сухожильного коленного рефлекса, который проявляется в разгибании в коленном суставе при лёгком постукивании по сухожилию ниже коленной чашечки (рис. 15, А).
Путь, по которому проходит нервный импульс при осуществлении рефлекса, называют рефлекторной дугой. В самые простые рефлекторные дуги входят только по два нейрона, в более сложные – по три, а в большинстве рефлекторных дуг насчитывается ещё больше нейронов. Примером двухнейронной рефлекторной дуги является дуга сухожильного коленного рефлекса, который проявляется в разгибании в коленном суставе при лёгком постукивании по сухожилию ниже коленной чашечки (рис. 15, А).
В состав трёхнейронной рефлекторной дуги входят: 1) рецептор; 2) афферентный нейрон; 3) вставочный нейрон; 4) эфферентный нейрон; 5) рабочий орган (клетки мышцы или железы). Связь между нейронами в рефлекторной дуге, между эфферентным нейроном и клетками рабочего органа осуществляется с помощью синапсов.
Рецепторами называют окончания дендритов афферентных нейронов, а также специализированные клетки (например, палочки и колбочки сетчатки глаза), которые воспринимают раздражение и в ответ на него генерируют нервные импульсы. Нервные импульсы от рецептора поступают по афферентному нервному пути, состоящему из дендрита, тела и аксона афферентного нейрона, в нервный центр.
Нервным центром называют совокупность нейронов, необходимых для осуществления рефлекса или регуляции той или иной функции. Большинство нервных центров находится в ЦНС, но они также есть и в нервных узлах периферической нервной системы. В один нервный центр могут функционально объединяться нейроны, тела которых лежат в разных отделах нервной системы.
В нервном центре расположен вставочный нейрон, на тело или дендриты которого передаётся возбуждение с аксона афферентного нейрона. По аксону вставочного нейрона импульс поступает к эфферентному нейрону, тело которого тоже находится в нервном центре. В большинстве рефлекторных дуг между аксоном афферентного нейрона и телом эфферентного нейрона включается не один, а целая цепь вставочных нейронов. Такие рефлекторные дуги называют полинейронными, или полисинаптическими.
По аксону эфферентного нейрона нервные импульсы поступают к клеткам рабочего органа (мышцы, железы). В результате наблюдается рефлекторная реакция (движение, выделение секрета) на раздражение рецепторов. Время от начала раздражения рецепторов до начала ответной реакции называют временем реакции, или латентным временем рефлекса. Больше всего время рефлекса зависит от скорости проведения возбуждения через нервные центры. Ухудшение функционального состояния нервного центра приводит к увеличению времени рефлекса.
Выполнение ответной реакции ещё не является окончанием рефлекторного акта. В осуществляющем ответную реакцию рабочем органе раздражаются рецепторы, импульсы от которых поступают по афферентным нервным волокнам в ЦНС и информируют нервные центры о протекании рефлекторной реакции и состоянии рабочего органа. Такую информацию называют обратной связью. Различают положительные и отрицательные обратные связи. Положительные обратные связи вызывают продолжение и усиление ответной рефлекторной реакции, а отрицательные обратные связи – её ослабление и прекращение.
Таким образом, возбуждение при рефлекторной реакции не только передаётся по рефлекторной дуге от первоначально раздражаемого рецептора к рабочему органу, но и затем снова поступает в ЦНС от рецепторов рабочего органа, которые возбудились в результате его ответной рефлекторной реакции. Такая взаимосвязь между нервными центрами и иннервируемыми органами, которая наблюдается при осуществлении рефлекса, называется рефлекторным кольцом. Благодаря обратным связям, осуществляющимся по рефлекторному кольцу, ЦНС получает информацию о результатах рефлекторных реакций, вносит поправки в их осуществление, обеспечивает координированную деятельность организма.
СИНАПТИЧЕСКИЕ СВЯЗИ И НЕЙРОННЫЕ ЦЕПИ
Межнейронные синаптические связи
Нервная система состоит из отдельных нейронов и поэтому нуждается в механизмах, обеспечивающих межклеточные взаимодействия. Главная задача межнейронных связей заключается в передаче и обработке информации, закодированной в виде электрических сигналов – нервных импульсов. Передача импульсов с нейрона на нейрон происходит в синапсах, которые по механизму передачи могут быть электрическими, химическими и очень редко смешанными. Синапсы с электрическим механизмом передачи чаще встречаются у животных с более примитивной нервной системой, хотя они и обнаружены в мозге млекопитающих, включая приматов. Их число уменьшается в процессе эмбрионального развития. Синапсы с химическим механизмом передачи составляют большую часть синаптического аппарата центральной нервной системы высших животных и человека.
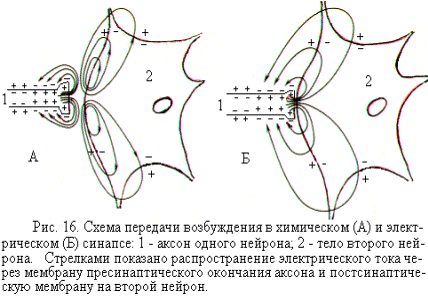 В электрических синапсах ширина синаптической щели значительно меньше, чем в химических синапсах и составляет всего 2 – 4 нм. Через синаптическую щель перекинуты белковые мостики, которые представляют собой своеобразные каналы, пронизывающие пре- и постсинаптическую мембраны синапса. По этим каналам могут переходить из нейрона в нейрон ионы и даже небольшие молекулы, благодаря чему электрическое сопротивление в области такого синапса оказывается очень низким. Это позволяет пресинаптическому току (нервному импульсу) распространяться на постсинаптическую клетку примерно так же, как происходит проведение нервного импульса по нервному волокну (рис. 16). Таким образом, в электрическом синапсе генератор постсинаптического тока находится в пресинаптической мембране, где возникает потенциал действия. Из неё он пассивно электротоническим способом распространяется на мембрану постсинаптического нейрона.
В электрических синапсах ширина синаптической щели значительно меньше, чем в химических синапсах и составляет всего 2 – 4 нм. Через синаптическую щель перекинуты белковые мостики, которые представляют собой своеобразные каналы, пронизывающие пре- и постсинаптическую мембраны синапса. По этим каналам могут переходить из нейрона в нейрон ионы и даже небольшие молекулы, благодаря чему электрическое сопротивление в области такого синапса оказывается очень низким. Это позволяет пресинаптическому току (нервному импульсу) распространяться на постсинаптическую клетку примерно так же, как происходит проведение нервного импульса по нервному волокну (рис. 16). Таким образом, в электрическом синапсе генератор постсинаптического тока находится в пресинаптической мембране, где возникает потенциал действия. Из неё он пассивно электротоническим способом распространяется на мембрану постсинаптического нейрона.
В химических синапсах синаптическая щель относительно широка (20 – 50 нм), а синаптические мембраны имеют высокое электрическое сопротивление. Поступающий в пресинаптический отдел нервный импульс представляет собой ток, который выходит через пресинаптическую мембрану и почти целиком уходит через широкую синаптическую щель мимо постсинаптической клетки. На постсинаптическую мембрану попадает незначительно малая часть этого тока, которая не способна вызвать заметный сдвиг мембранного потенциала постсинаптического нейрона. Поэтому в химических синапсах мембранный потенциал постсинаптиче-ского нейрона изменяется с помощью специальных химических веществ – медиаторов.
Нервный импульс, поступающий в пресинаптический отдел, вызывает вход в него ионов кальция. Это приводит к выбросу медиатора из синаптических пузырьков в синаптическую щель. Воздействуя на постсинаптические рецепторы, медиатор способен изменять состояние ионных каналов постсинаптической мембраны. Изменение ионной проницаемости постсинаптической мембраны, в свою очередь, вызывает её деполяризацию – возбуждающий постсинаптический потенциал. Таким образом, в химическом синапсе генератор постсинаптического тока находится непосредственно в постсинаптической мембране и запускается медиатором, выделяемым пресинаптическим окончанием.
|
|
|
|
|
Дата добавления: 2014-12-26; Просмотров: 2231; Нарушение авторских прав?; Мы поможем в написании вашей работы!