КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Голосеменные: их характеристика и основные представители
Выход растений на сушу как важнейший этап в развитии растительного мира. Особенности жизни растений в наземных условиях, приспособления растений к наземному образу жизни. Первые наземные растения. Особенности их строения и биологии.
Сравнительная характеристика голосеменных и покрытосеменных растений
| № | ОТЛИЧИТЕЛЬНЫЕ ПРИЗНАКИ | ОТДЕЛ ГОЛОСЕМЕННЫЕ(Gymnospermatophyta) | ОТДЕЛ ЦВЕТКОВЫЕ (Angiospermae) |
| Происхождение названия | Семечко сидит открыто на семенной чешуе («голо» + «семечка») | Семечко находится внутри плода («покрыто» + «семечка») | |
| Количество видов | Около 800 | Не менее 250 000 | |
| Филогенез | Появились в конце девонского периода | Появились в юрском периоде | |
| Жизненные формы | Деревья, иногда кустарники | Деревья, кусты, травы (одно-, двух-и многолетние) | |
| Элементы проводящей ткани | В большинстве нет сосудов, ситоподобные трубки без клеток-спутниц (но есть альбуминоидные клетки, которые выполняют ту же функцию, что и клетки-спутницы) | Есть сосуды, ситоподобные трубки с клетками-спутницами | |
| Луковицы | Отсутствуют | Есть | |
| Клубни | Отсутствуют | Есть | |
| Корневища | Отсутствуют | Есть | |
| Семенные зачатки | Расположенные открыто на мегаспорофилах | Находяца под покровом плодолистиков пестика | |
| Женский гаметофит (мегагаметофит) | Эндосперм с двумя или более архегониям | 8-ядерный зародышевый мешок (состоит из 7 клеток) | |
| Мужской гаметофит (микрогаметофит) | Пыльца, состоящая из двух проталиальних клеток, генеративной и сифоногенных клеток | Пыльца, состоящая из 2 клеток: сифоногенной и генеративной | |
| Опыление | С помощью ветра | С помощью ветра, воды, насекомых, птиц, млекопитающих т.д. | |
| Оплодотворение | Оплодотворение простое: один из спермиев сливается с яйцеклеткой одного из архегоний, проходит очень медленно (до 13 месяцев) | Оплодотворение двойное: один спермий сливается с яйцеклеткой, а другой — с ядром центральной клетки (2n) Происходит быстро | |
| Образование семян | Происходит очень медленно (до 1,5 — 2 лет) | Происходит сравнительно быстро — от 3-4 недель до одного вегетационного периода | |
| Эндосперм | Гаплоидный (п) | Триплоидный (Зп) | |
| Зародыш | Имеет две семядоли (до 15) | Имеет 1-2 семядоли | |
| Семена | Расположены в шишках, сидят открыто («голо») на семенных чешуйках | Находятся внутри плода (покрыто околоплодником) | |
| Значение в эволюции животных | Не могли создать достаточную кормовую базу для птиц и млекопитающих | Благодаря возникновению плода обеспечили достаточную кормовую базу для млекопитающих и птиц |
Появление наземных, или высших, растений ознаменовало начало новой эры в жизни нашей планеты. Освоение растениями суши сопровождалось появлением новых, наземных, форм животных; сопряженная эволюция растений и животных привела к колоссальному разнообразию жизни на земле, изменила ее облик. Первые достоверные наземные растения, известные только по спорам, датируются началом силурийского периода. Из верхнесилурийских и нижнедевонских отложений по сохранившимся макроостаткам или по отпечаткам органов описаны наземные растения. Эти первые известные нам высшие растения объединены в группу риниофитов. Несмотря на анатомическую и морфологическую простоту строения, это были уже типичные наземные растения. Об этом свидетельствует наличие кутинизированной эпидермы с устьицами, развитой водопроводящей системы, состоящей из трахеид, и наличие многоклеточных спорангиев с кутинизированными спорами. Следовательно, можно предположить, что процесс освоения суши растениями начался значительно раньше - в кембрии или в ордовике.
Предпосылок для появления наземных растений было, по-видимому, несколько. Во-первых, независимый ход эволюции растительного мира подготовил появление новых, более совершенных форм. Во-вторых, за счет фотосинтеза морских водорослей в атмосфере земли произошло увеличение количества кислорода; к началу силурийского периода оно достигло такой концентрации, при которой оказалась возможной жизнь на суше. В-третьих, в начале палеозойской эры на обширных территориях земли происходили крупнейшие горообразовательные процессы, в результате которых возникли Скандинавские горы, горы Тянь-Шань, Саяны. Это вызвало обмеление многих морей и постепенное появление суши на месте бывших мелких водоемов. Если раньше водоросли, населявшие литоральную зону, только в отдельные кратковременные периоды жизни оказывались вне воды, то по мере обмеления морей они переходили к более длительному пребыванию на суше. Это, очевидно, сопровождалось массовой гибелью водорослей; выживали лишь те немногие растения, которые смогли противостоять новым условиям жизни.
В ходе длительного эволюционного процесса возникали новые виды, постепенно формировавшие типичные наземные растения.
К сожалению, промежуточные формы палеонтологическая летопись не сохранила. Новая воздушно-наземная среда обитания оказалась чрезвычайно противоречивой, принципиально отличной от первоначальной водной. Прежде всего она характеризовалась повышенной солнечной радиацией, дефицитом влаги и сложными контрастами двухфазной воздушно-наземной среды. Вполне вероятно допустить, что у некоторых переходных форм в процессе обмена веществ мог вырабатываться кутин, который откладывался, на поверхности растений. Это было первым этапом формирования эпидермы. Чрезмерное выделение кутина неминуемо приводило к гибели растений, так как сплошная пленка кутина препятствовала газообмену. Только те растения, у которых выделялось умеренное количество кутина, смогли сформировать сложную специализированную ткань - эпидерму с устьицами, способную и защищать растение от высыхания, и осуществлять газообмен. Таким образом, важнейшей тканью наземных растений, без которой невозможно освоение суши, следует считать эпидерму. Однако возникновение эпидермы лишало наземные растения возможности поглощать воду всей поверхностью, как это происходит у водорослей.
У самых первых наземных растений, имевших еще небольшие размеры, поглощение воды осуществлялось с помощью ризоидов - одноклеточных или многоклеточных однорядных нитей. Однако по мере увеличения размеров тела происходил процесс формирования сложных специализированных органов - корней с корневыми волосками. По-видимому, образование корней, начавшееся с верхнедевонского периода, в разных систематических группах растений происходило разными путями.
Происхождение растений связано с первыми этапами развития жизни на Земле. Разнообразный и богатый мир растений возник за многие миллионы лет в ходе длительной эволюции. Менялись условия жизни на планете, вслед за ними менялись и растения. Первоначально жизнь зародилась в воде. До сих пор в океанах можно найти примитивные одноклеточные и колониальные растения: сине-зеленые и зеленые водоросли.
Организмы, похожие на них появились в архейскую эру (более 3 млрд. лет назад). В протерозойскую эру (2 млрд лет назад) определились две линии развития жизни животных и растений: водная и сухопутная; в это время и возникли сине-зеленые водоросли и некоторые настоящие водоросли. Когда растения стали завоевывать сушу - неизвестно; первые микроскопические наземные растения появились на границе протерозоя и палеозоя, около 420 млн лег назад. Но как только растения вышли на сушу темпы их эволюции сильно ускорились. Ученые полагают, что все наземные растения, существующие сейчас, произошли от риниофитов (это тип споровых растений, представленный самыми примитивными сосудистыми формами).
В девонском периоде палеозойской эры (около 416 млн. лет назад) растения уже заселили сушу. Это были древовидные папоротники, хвощи, древние голосеменные, а также растения, не имеющие корней и селящиеся на других

Все растения на планете произошли от споровых растений ринофитов. Рисунок из книги Э. Геккеля

Сосна, голосеменное (хвойное) растение деревьях и растениях. Пышная растительность изменила состав атмосферы, обогатила ее кислородом. Благодаря отмершим растениям увеличился слой почв, произошло и усложнение животных организмов. У растений развились проводящая и корневая системы. Господствующее положение в палеозое заняли голосеменные (или хвойные) растения, у которых оплодотворение происходило без участия воды, а зародыш был защищен от неблагоприятных условий, в семени находился запас питательных веществ, необходимых для развития зародыша.
В мезозойскую эру (примерно 140 млн лет назад) возникли покрытосеменные, или цветковые растения, ставшие затем господствующими на нашей планете. Но в те далекие времена привычные нам цветковые растения имели древовидную форму: это были гигантские магнолии, лавры, платаны, эвкалипты, ивы, виноград, тополя, древовидные пальмы. Травянистая растительность была развита значительно меньше.
В кайнозойскую эру (а мы с вами живем в антропогенный период кайнозойской эры) цветковые растения достигли наивысшего развития, и постепенно увеличили территории своего распространения. В этот период формировалась современная зональность растительности, т. е. растения приспособились к жизни в разных климатических поясах, от экватора до полярных широт. В истории развития растительного мира одним из важнейшие моментов является возникновение сухопутной флоры. В то время как бактерии и водоросли живут или в водоемах, или, если они обитают в почве, на сырых склонах или на голой земле, в очень тонком слое воды, достаточном для свободного их передвижения, так как сами они очень малы, высшие растения — мхи, папоротникообразные, голосеменные и цветковые, образующие ныне главную массу земной (растительности, — приводят в соприкосновение с водой только свои корни, корневые волоски и ризоиды, все же остальные их органы находятся в сухой атмосфере и испаряют воду всей поверхностью.
Если предположить, что родоначальниками растений суши были водоросли, то трудно понять, каким образом эти легко гибнущие на воздухе организмы могли приспособиться к резкой потере воды и ее возмещению.
Познакомимся вкратце с теорией Бауера, взявшегося выяснить этот трудный вопрос. Основой всякого размножения является деление клеточного ядра, влекущее за собой и деление клетки. Если организм одноклеточный, то посло деления клетки получается два организма. Если организм сложный, то в результате деления получается рост ткани, а рост ткани влечет за собой и рост органа, в состав которого входит данная ткань. Само собой разумеется, что каждая клетка при делении дает себе подобную.
Обратим теперь внимание на строение ядра из нитей особого ядерного вещества, или хроматина. В период деления хроматин ядра распределяется между определенным числом отрезков, называемых хромосомами. Согласно данным науки о наследственности, каждая хромосома имеет особое значение как носитель определенных наследственных свойств организма. При передаче этих свойств от одного поколения другому, передача совершается именно через хромосомы ядра. Если одна из хромосом погибнет, то и свойства, носителем которых она является, не передадутся по наследству. Так вот по отношению к хромосомам ядра одного и того же организма неодинаковы. Как правило можно принять, что число хромосом у каждого обособленного организма свое, постоянное. Так, капуста имеет в своих клетка 18 хромосом, рапс 20, у табака и махорки их 24, у липы 41. у осины 8, у одних сортов хлопка 26, у других 52, у кукурузы 20, у пшеницы 42, у ржи 14 и так далее. При делении ядра. в стадии экваториальной пластинки, хромосомы делятся каждая продольно на две, после чего начинают расходиться к полюсам старого ядра, образуя дочерние ядра. Поэтому-то в каждом дочернем ядре будет то же число хромосом, что было в материнском ядре, а самое деление называется уравнительным или эквационным. Однако после неопределенно большого числа эквационных делений клетки начинают делиться иначе. Именно, перед образованием ядерного веретена хромосомы попарно сливаются, и число их уменьшается вдвое. В стадии ядерной пластинки они снова делятся продольно и расходятся, образуя два дочерних ядра, но так как слияние уменьшило их число вдвое, то и дочерние ядра будут обладать лишь половинным числом хромосом. Полное число хромосом называется диплоидным, или двойным, а половинное — гаплоидным, или простым. Из гаплоидной клетки не может развиться такой же организм, как из диплоидной, а будет развиваться организм более слабый, в котором число хромосом все время будет поддерживаться эквационным делением все в том же гаплоидном числе. Только слияние двух ядер в процессе оплодотворения удваивает число хромосом, и возникающий в результате молодой организм снова становится диплоидным. Диплоидный организм размножается обычно почкованием. Если почки одноклеточные, то они называются спорами, если состоят из однородной ткани, то таллидиями, если из органов, каковы стебель и листья, то просто почками. Гаплоид может размножаться подобно диплоиду, оставаясь при этом подобным себе самому, но может и дать так называемые гаметы, т. е. клетки, не способные к самостоятельному развитию, но способные к слиянию друг с другом, после чего число хромосом удваивается и получается так называемая зигота, т. е. двойная клетка, первая клетка следующего поколения. У многих растений, как, например, у мхов и папоротников, у многих водорослей поколение диплоидных клеток неизбежно сменяется поколением гаплоидных. У мхов сам мох гаплоидный, диплоидным же является развивающийся на нем спорогон, состоящий из пяты, ножки, апофиза и коробочки; у папоротников сам папоротник, приносящий во множестве споры, является диплоидным, а вырастающие из спор маленькие заростки гаплоидны.
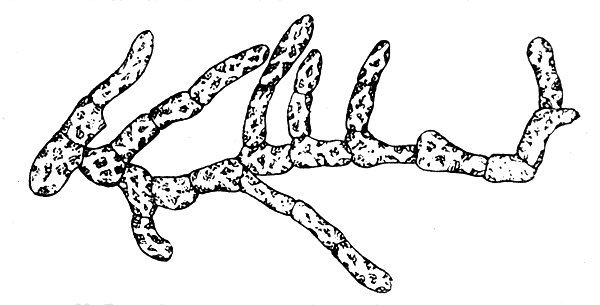
Рис. 22. Гаметофит, или заросток (гаплоид) водоросли ламинария. Сильно увеличено
Как правило можно вывести из этих я многих других примеров, что диплоид приносит посоле наступления в некоторых его клетках редукционного деления споры, а из cпор вырастают гаплоидные организмы, заканчивающие свою жизнь развитием гамет. Гаметы сливаются, удваивая это число хромосом и образуя зиготу, из которой вырастает диплоидный организм. Два типа размножения: почкование и слияние гамет соответствуют как бы двум различным по своей внешности и свойствам организмам, следующим в обязательном порядке один за другим. Это явление носит в науке наименование смены поколений, или смены спорофита гаметофитом.
Поразительна смена гаметофита спорофитом у морской водоросли ламинарии.
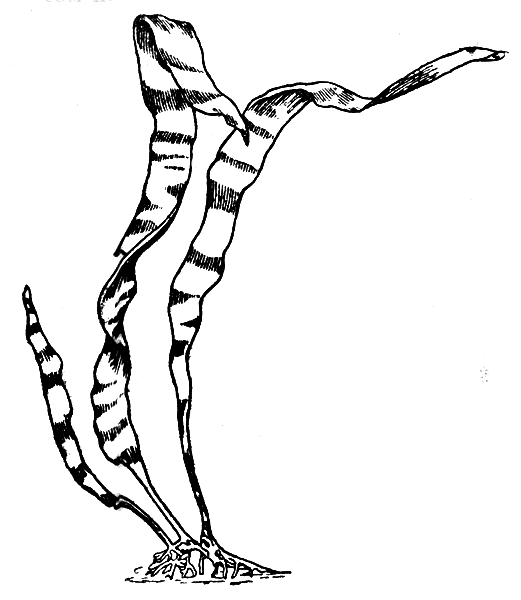
Рис. 23. Спорофит (диплоид) водоросли ламинария. Сильно уменьшено
Несмотря на широкое географическое распространение, крупные размеры и большое промысловое значение этой водоросли, вплоть до работ Соважо (1915) и Килина (1916), ботаники считали ламинарию, или морскую капусту, за организм, не имеющий полового размножения, так как известны были только ее подвижные споры, гаметы же никогда не наблюдались. Оказалось, однако, и притом совершенно неожиданно, что споры ламинарии прикрепляются к камням и разрастаются в маленькие растеньица, которые после продолжительного периода развития дают: одни — яйцевые клеточки, другие — подвижные сперматозоиды. Оплодотворенная яйцеклетка разрастается в крупное растение с дифференцированными тканями, приносящее затем периодически массу спор.
Для спорофита-диплоида ламинарии характерны быстрый рост, мощность развития и физиологическое расчленение тканей тела на покровные, ассимиляционные, проводящие, механические и выделительные; для гаметофита гаплоида (20 хромосом) — малый рост, медленное развитие, простота строения.
За исключением мхов у всех растений, у которых ясно выражена смена поколений, мы находим то же взаимоотношение, т. е. слабый и просто организованный, медленно растущий гамеофит, хорошо развивающийся только в воде или при ее изобилии, и быстро растущий, сложно построенный и способный сопротивляться высыханию спорофит. Ясно, что обладание двойным числом хромосом, короче, двойным ядром, чрезвычайно выгодно для организма. Ясно также, что естественный отбор должен давать в борьбе за существование преобладание тем растениям, диплоидный аппарат которых развит сильнее. Если в водной среде гаплоидные организмы и могут сравнительно счастливо заканчивать свой жизненный цикл, то на суше солнце и ветер быстро сокращают их жизнь. Но в воде существование гаплоидных организмов поддерживается только напряженностью их размножения, при полной нестойкости особи. Диплоид не таков — надо видеть, как рвет и треплет океанский прибой гибкое тело ламинарии, растущей близ берега на подводных скалах, чтобы оценить всю силу ее сопотивляемости.
Выше мы видели, что первый расцвет растительной жизни на суше был приурочен к приморским местностям, особенно к берегам лагунных озер и болот, лежащих между песчаными береговыми валами и подгорной террасой. Здесь впервые выработался тип растения, нижней своей частью находящегося в воде, а верхушкой в воздушной среде под прямыми лучами солнца. Споры, развивавшиеся в мелкой нагретой воде лагун, при понижении уровня воды, вследствие испарения, образовывали заростки в виде плоских зеленых лопастных пластинок. Отдельные участки их, выдвигаясь из воды и испаряя воду, вызывали возникновение токов воды, возмещавших испаряемую, и этим способствовали росту своих тканей. Образовывалось подобие первичного стебля. Если на таком стебле возникали споры или гаметы, то они падали в воду и там прорастали, образуя новые заростки, или же, сливаясь в воде, облегчавшей их движения, давали зиготы, полагавшие начало диплоидному поколению — спорофиту.
Позднее, проникновением на незатопляемую сушу, первобытные растения развили корень (часть спорофита) и получили способность использовать грунтовые воды, что дает возможность переживать промежутки между дождями. Осталось только выработать такой способ образования гамет и их слияния, который за неимением у корней данного растения свободной воды, позволял бы гаметам двигаться во внутренней среде самого растения. Теперь почти все клетки сложного растительного организма становятся диплоидными, почему оно приобретает и быстроту развития, и способность к расчленению клеток на ткани, способность к выработке защитных приспособлений, каких не знали его жившие у воды предки. Гаплоидными остаются только споры и вырастающие из них микроскопические заростки, защищенные тканями тела растения матери. Таким образом, выход растений на сушу сопровождался полным подчинением гаметофита спорофиту и сильным разрастанием последнего.
Первые голосеменные появились в конце девонского периода около 350 млн. лет назад; вероятно, они произошли от древних папоротниковидных, вымерших в начале каменноугольного периода. В мезозойскую эру - эпоху горообразования, поднятия материков и иссушения климата - голосеменные достигли расцвета, но уже с середины мелового периода уступили свое господствующее положение покрытосеменным.
Отдел современных голосеменных насчитывает более 700 видов. Несмотря на относительно малую численность видов, голосеменные завоевали почти весь земной шар. В умеренных широтах Северного полушария они на огромных пространствах образуют хвойные леса, называемые тайгой.
Сейчас на Земле произрастают представители 4 классов голосеменных. Из них Гинкговые (единственный вид - Ginkgo biloba) и Саговниковые (120-130 видов 10 тропических родов) - это остатки процветавших в мезозое групп. Возраст и происхождение Гнетовых (71 вид 3 родов) неясны из-за отсутствия палеоботанических данных, и лишь класс Хвойных (Pinopsida) широко представлен ныне на нашей планете, по своему значению превосходя все остальные группы высших растений. В настоящее время насчитывается не менее 560 видов из 55 родов хвойных, распространенных в умеренных областях обоих полушарий. Подавляющее большинство представителей этого класса - вечнозеленые древесные растения, возраст и размеры которых потрясают воображение. Так, известны экземпляры секвойи вечнозеленой высотой более 100 м и диаметром ствола до 11 м. Некоторые экземпляры мамонтова дерева достигают диаметра 12 м, а таксодиума мексиканского - 16 м. Рекорд долгожительства принадлежит североамериканской сосне долговечной - до 4900 лет! Кроме деревьев, широко встречаются кустарниковые и даже кустарничковые хвойные (новозеландская сосна-пигмей).
Древняя группа (отдел) семенных растений, занимающая промежуточное положение между папоротниками и покрытосеменными (цветковыми) растениями. От папоротников отличаются наличием семезачатков (семяпочек), от покрытосеменных - отсутствием плодолистиков. Современные голосеменные представлены преимущественно деревьями, значительно реже - кустарниками и очень редко - лианами; травянистых растений среди них нет. Листья голосеменных значительно отличаются от других групп растений не только по форме и размерам, но и по морфологии и анатомии. У большинства видов они игловидные (хвоя) или чешуевидные; у отдельных представителей они крупные (например, у вельвичии удивительной их длина достигает 2-3 м), перисторассеченные, двулопастные и др. Листья располагаются поодиночке, по два или несколько в пучках.
Все современные виды голосеменных имеют стробилы - укороченные и ограниченные в росте репродуктивные побеги. Большинство голосеменных - деревья или кустарники, нередко достигающие огромных размеров. Некоторые сильно разветвлены и несут много небольших или мелких, часто чешуевидных листьев (кордаитовые, гинкговые и хвойные), другие не ветвятся или ветвятся очень слабо и имеют относительно крупные, большей частью перистые, листья (лигиноптеридовые, беннеттитовые, цикадовые и вельвичия). Листья сильно различаются не только по числу и размерам, но и по анатомическому строению. Водопроводящая система (ксилема) голосеменных состоит преимущественно из трахеид, и лишь у некоторых групп имеются настоящие сосуды.
Подавляющее большинство голосеменных - вечнозеленые, одно - или двудомные растения с хорошо развитыми стеблем и корневой системой, образованной главным и боковым корнями. Расселяются они семенами, которые формируются из семязачатков. Семязачатки голые (отсюда название отдела), расположены на мегаспорофиллах или на семенных чешуях, собранных в женские шишки.
В цикле развития голосеменных наблюдается последовательная смена двух поколений - спорофита и гаметофита с господством спорофита. Гаметофиты сильно редуцированы, причем мужские гаметофиты голо - и покрытосеменных растений не имеют антеридиев, чем резко отличаются от всех разноспоровых бессеменных растений.
Голосеменные включают шесть классов, два из которых полностью исчезли, а остальные представлены ныне живущими растениями. Наиболее сохранившейся и самой многочисленной группой голосеменных является класс Хвойные, насчитывающий не менее 560 видов, образующих леса на обширных пространствах Северной Евразии и Северной Америки. Наибольшее число видов сосны, ели, лиственницы встречается у побережий Тихого океана.
Класс Хвойные
Все хвойные - вечнозеленые, реже листопадные (например, лиственница) деревья или кустарники с игольчатыми или чешуевидными (например, у кипариса) листьями. Игловидные листья (хвоя) плотные, кожистые и жесткие, покрыты толстым слоем кутикулы. Устьица погружены в углубления, заполненные воском. Все эти особенности строения листьев обеспечивают хорошее приспособление хвойных к произрастанию как в засушливых, так и в холодных местообитаниях.
Сосна обыкновенная - однодомное растение. В мае у основания молодых побегов сосны образуются пучки зеленовато-желтых мужских шишек длиной 4-6 мм и диаметром 3-4 мм. На оси такой шишки расположены многослойные чешуйчатые листочки, или микроспорофиллы. На нижней поверхности микроспорофиллов находятся два микроспорангия - пыльцевых мешка, в которых образуется пыльца. Каждое пыльцевое зерно снабжено двумя воздушными мешками, что облегчает перенос пыльцы ветром. В пыльцевом зерне имеются две клетки, одна из которых впоследствии, при попадании на семязачаток, формирует пыльцевую трубку, другая после деления образует два спермия.
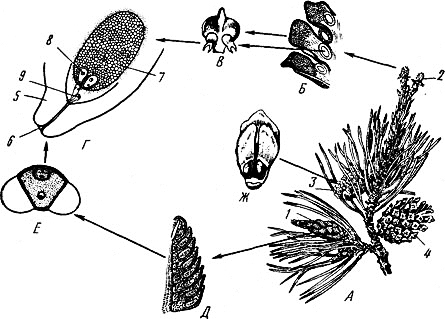
Рис.1. Цикл развития сосны обыкновенной
а - ветка с шишками; б - женская шишка в разрезе; в - семенная чешуя с семязачатками; г - семязачаток в разрезе; д - мужская шишка в разрезе; е - пыльца; ж - семенная чешуя с семенами; 1 - мужская шишка; 2 - молодая женская шишка; 3 - шишка с семенами; 4 - шишка после высыпания семян; 5 - пыльцевход; 6 - покров; 7 - пыльцевая трубка со спермиями; 8 - архегоний с яйцеклеткой; 9 - эндосперм
На других побегах того же растения образуются женские шишки красноватого цвета. На их главной оси располагаются мелкие прозрачные кроющие чешуйки, в пазухах которых сидят крупные толстые, впоследствии одревесневающие чешуи. На верхней стороне этих чешуй расположено по два семязачатка, в каждом из которых развивается женский гаметофит - эндосперм с двумя архегониями с крупной яйцеклеткой в каждом из них. На верхушке семязачатка, снаружи защищенного интегументом, имеется отверстие - пыльцевход, или микропиле.
Поздней весной или в начале лета созревшая пыльца разносится ветром и попадает на семязачаток. Через микропиле пыльца втягивается внутрь семязачатка, где и прорастает в пыльцевую трубку, которая проникает к архегониям. Образовавшиеся к этому времени два спермия по пыльцевой трубке попадают к архегониям. Затем один из спермиев сливается с яйцеклеткой, а другой отмирает. Из оплодотворенной яйцеклетки (зиготы) формируется зародыш семени, а семязачаток превращается в семя. Семена у сосны созревают на второй год, высыпаются из шишек и, подхваченные животными или ветром, переносятся на значительные расстояния.
По своему значению в биосфере и роли в хозяйственной деятельности человека хвойные занимают второе место после покрытосеменных, далеко превосходя все остальные группы высших растений. Они помогают решать огромные водоохранные и ландшафтные задачи, служат важнейшим источником древесины, сырья для получения канифоли, скипидара, спирта, бальзамов, эфирных масел для парфюмерной промышленности, лекарственных и других ценных веществ. Некоторые хвойные культивируются как декоративные (пихты, туи, кипарисы, кедры и др.). Семена ряда сосен (сибирской, корейской, итальянской) употребляются в пищу, из них также получают масло.
Представители других классов голосеменных (саговниковые, гнетовые, гинкговые) встречаются значительно реже и менее известны, чем хвойные. Однако почти все виды саговниковых декоративны и пользуются широкой популярностью у садовников многих стран. Вечнозеленые безлистные невысокие кустарники эфедры (класс гнетовых) служат источником сырья для получения алкалоида эфедрина, который применяется как средство, возбуждающее центральную нервную систему, а также при лечении заболеваний аллергического характера.
|
|
Дата добавления: 2015-04-25; Просмотров: 2239; Нарушение авторских прав?; Мы поможем в написании вашей работы!