КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Отдел Гетероконтные (разножгутиковые) водоросли – heterokontophyta 2 страница
|
|
|
|
Порядок Хромулиновые – Chromulinales. К этому порядку относят организмы с монадным, пальмеллоидным и амебоидным типами дифференциации таллома. Монадные клетки с одним жгутиком, видимым в световой микроскоп.
Род Хризамеба (рис. 37, А) – пресноводные амебоидные водоросли, они имеют в жизненном цикле жгутиковую стадию. Зигота превращается в стоматоцисту.
Род Хромулина (рис. 37, В) - одноклеточные свободноплавающие организмы, обитающие преимущественно в пресных водах. Клетки голые; имеется одна или две золотистые пластиды.
Род Динобрион (рис. 37, Г) – одноклеточные и колониальные, свободноплавающие или прикрепленные представители. Монады находятся в вазообразных домиках, образованных целлюлозными микрофибриллами. Помимо целлюлозы в состав домика входит и большое количество аминокислот. Кустикообразная форма колоний связана со способом вегетативного деления клеток, когда одна из дочерних клеток, покидая родительский домик, прикрепляется к его отверстию. Вращаясь вокруг своей оси, она формирует собственный домик. Родительский домик могут покинуть и обе дочерние клетки. Клетки содержат 1-2 хлоропласта, на переднем конце клетки видны глазок и 2 сократительные вакуоли. Хризоламинарин расположен в вакуоли на заднем конце клетки. Размножение вегетативное и половое.
Порядок Гиббердиевые – Hibberdiales. Для представителей данного порядка характерен уникальный пигментный состав. Помимо фукоксантина они имеют дополнительный светособирающий каротиноидный пигмент – антераксантин.
Включает роды Гиббердия, Хромофитон, Стилоцерас, Хризопиксис, Платитека и другие.
Род Гиббердия имеет две стадии в жизненном цикле: колониальную пальмеллоидную неподвижную и одноклеточную монадную подвижную. При световой микроскопии у монадной стадии виден только один жгутик. Характерно наличие помимо фукоксантина еще одного дополнительного светособирающего каротиноидного пигмента – антераксантина.
|
|
|
Порядок Гидрурусовые – Hydrurales. Представители данного порядка имеют таллом с пальмеллоидным и псевдопаренхиматозным типами строения. Характерно наличие уникальных зооспор тетраэдрической формы. Глазка нет. Митоз полузакрытый.
Включает роды Гидрурус, Целлониелла, Федерматиум, Хризонебула и другие.
Род Гидрурус (рис. 37, Е) является наиболее дифференцированным среди пальмеллоидных водорослей. Его таллом представляет собой крупные, длиной до 30 см слизистые колонии, имеющие вид коричневых разветвленных шнуров, часто издающих очень неприятный запах. В колониях можно различить главный ствол и боковые разветвления. В клетках, погруженных в общую слизь, содержится один чашевидный хроматофор и несколько сократительных вакуолей. Клетки рыхло расположены по периферии колонии и более плотно в середине. Таллом может расти только за счет деления апикальных клеток. При бесполом размножении в клетках боковых ветвей колонии формируются зооспоры. Образует шаровидные цисты. Встречается в горных ручьях и реках с холодной водой, где прикрепляется к твердым субстратам.
Класс Синуровые водоросли – Sinurophyceae
Название класса происходит от греч. syn – вместе и ura – хвост. Эта группа организмов в 1987 г. выделена из золотистых водорослей. Объединяет монадные одиночные и колониальные организмы, иногда с чередованием в жизненном цикле монадной и пальмеллоидной стадии. Повержность пектиновых оболочек покрыта панцирем из кремниевых чешуек. Митохондрии с трубчатыми кристами, обычно расположены в цитоплазме вокруг хлоропласта. Ядро одно, хлоропластов, окруженных четырьмя мембранами, обычно два. Ламеллы трехтилакоидные, опоясывающая ламелла имеется. Основные пигменты – хлорофиллы а и с, β -каротин и фукоксантин. Стигмы нет. Клетки обычно с 2 неравными жгутиками. Длинный перистый жгутик направлен вперед. Короткий гладкий жгутик, который иногда сильно редуцирован, направлен назад.
|
|
|
Клетки размножаются в основном продольным делением. Колонии, распадаясь, дают начало молодым колониям. У некоторых видов описан половой процесс в форме изогамии. При этом, в отличие от золотистых водорослей, слияние изогамет у синуровых водорослей происходит не на передних, а на их задних концах. Эндогенным путем образуются окремнелые цисты с порой, сходные с таковыми у хризофит.
Преимущественно планктонные формы, пальмеллоидные стадии входят в состав бентоса. Большинство видов синуровых водорослей - фототрофные пресноводные организмы.
Систематика.
Класс Синуровые водоросли включает 7 порядков: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales и Thallochrysidales.
Порядок Синуровые – Synurales. Монадные формы, пектиновые оболочки клеток обычно покрыты панцирем из кремнеземных чешуек, сцементированных органическим веществом в цельный футляр. Хлоропластов один или два.
Род Синура (рис. 38) -пресноводные монадные формы с двумя неравными жгутиками и двумя пристеночными хлоропластами. Ядро грушевидное, расположено в передней части клетки. Позади ядра имеется одна большая вакуоль, а в задней части клетки – несколько мелких сократительных вакуолей. Клетки покрыты чешуйками, как черепицей Размножение чаще всего вегетативное, для некоторых известен половой процесс. Образует колонии (рис. 39, А). Часто во всех клетках колонии возникают цисты. При массовом развитии Синура может придавать воде неприятный запах.
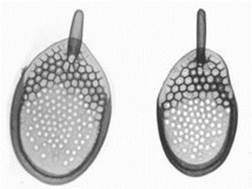
Рис. 38. Внешний вид клеток Синуры
Порядок Ризохлоридовые – Rhizochloridales. Преимущественно пресноводные организмы с ризоподиальным типом дифференциации таллома, обитающие главным образом в пресных водоемах.
Род Малломонас (рис. 39, Б-Г) – одноклеточная монада с одним хорошо заметным жгутиком и одним раздвоенным хлоропластом.

Рис. 39. Внешний вид синуровых водорослей (по: Г.А. Белякова и др., 2006): А – делящаяся колония Синуры; Б, В – образование статоспоры и Г – внешний вид Малломонаса
|
|
|
Клетка покрыта чешуйками черепитчато и по спирали, некоторые из них несут игловидные шипы (рис. 39, Г). Характерно наличие статоспор (рис. 39, Б, В). Для ряда представителей описан половой процесс - гологамия.
Род Миксохлорис обитает в клетках листьев сфагновых мхов, имеет вид крупного многоядерного плазмодия. Осенью он образует цисты, которые прорастают весной. Из них выходят зооспоры или амёбы, проникающие в пустые клетки листьев и сливающиеся там в плазмодий.
Порядок Хлорамёбовые – Chloramoebales. Включает монадных представителей. Встречаются в соленых и пресных водах.
Род Гетерохлорис – его клетки могут менять форму, образуя псевдоподии. Это явление присуще многим представителям порядка, так же как тенденция к животному способу питания. В клетке имеются несколько хлоропластов, капли масла и хризоламинарин. В передней части клетки имеются сократительные вакуоли, в центре - одно ядро. Размножается вегетативно продольным делением клетки.
Порядок Охромонадовые — Ochromonadales. Объединяет голые формы с двумя неравными жгутиками. Пресноводные и морские формы.
Род Охромонас включает одноклеточные монады с двумя неравными жгутиками. Клетки одеты только плазмалеммой. На переднем конце клетки есть сократительная вакуоль и глазок, на заднем конце — вакуоль с хризоламинарином. Размножается вегетативно, борозда деления начинается у переднего конца клетки между двумя парами жгутиковых оснований. Виды рода распространены в олиготрофных пресные водах, но имеются и морские представители.
Порядок Гетероглеевые — Heterogloeales. Включает водоросли с пальмеллоидным типом дифференциации таллома. Представители порядка встречаются чаще в пресных, чем в соленых водах.
Род Гельминтоглея обитает в солоноватых водах. Эта колония, сидящая на расширенном основании, состоит из ветвящихся слизистых тяжей. В этой слизи беспорядочно располагаются протопласты, каждый из которых окружен собственной слизистой обверткой.
Класс Феотамниевые водоросли – Phaeothamniophyceae
|
|
|
Название класса происходит от типового рода Phaeothamnion (от греч. phaeos – смуглый кустик). Представителей данной группы организмов выделили в отдельный класс из желтозеленых и золотистых водорослей в 1998 г. на основании анализа последовательности рибосомальных генов и особенностей ультраструктуры клетки. Для феотамниевых водорослей характерна уникальная комбинация пигментов: фукоксантина с гетероксантином, отсутствие виолаксантина.
Особенности строения клетки: отсутствие вакуолей с хризоламинарином и отсутствие эндогенных цист с кремнеземными стенками.
Класс включает одноклеточные, колониальные и многоклеточные организмы с коккоидным, пальмеллоидным и нитчатым типом структуры таллома. Жгутиковые стадии с двумя неравными жгутиками. Жгутики латеральные или субапикальные. Стигма встречается у зооспор. Пигменты – хлорофиллы а и с, β -каротин, фукоксантин, диадиноксантин, диатоксантин и гетероксантин. Основным запасным продуктом является парамилон (β -глюкан). Клеточная стенка имеется, во время клеточного деления дочерняя стенка формируется внутри родительской стенки.
У феотамниевых из основных способов размножения встречается лишь вегетативное и бесполое. Бесполое размножение осуществляется автоспорами или зооспорами. Половое размножение неизвестно.
Виды Феотамниевых водорослей обитают исключительно в пресных водах. Систематика.
В настоящее время подтверждена монофилетичность класса феотамниевых водорослей, и среди охрофит он более близок бурым и желтозеленым водорослям, чем золотистым. В настоящее время известно около 30 видов этих водорослей, относимых к одному порядку Феотамниевые – Phaeothamniales.
Род Феотамнион представляет собой прикрепленные ветвящиеся нити до 1 см высоты (рис. 40). Клетки цилиндрические, неправильные, расширенные кверху, по

Рис. 40. Внешний вид Феотамниона.
длине в два-три раза превышают ширину. При делении клеток материал родительской стенки расходуется на образование слоистого чехла, окружающего клетки нити. По периферии клетки под плазмалеммой имеются везикулы. Они напоминают физоды – образования, встречающиеся в клетках бурых водорослей. Клетки содержат от одной до нескольких дисковидных пластид оливково-бурого цвета. Зооспоры образуются по 1–2, реже по 4–8 в одной клетке. У зооспор жгутики прикрепляются сбоку. Положение жгутиковых корешков у Феотамниона напоминает таковое у желтозеленых и бурых водорослей. У него также отсутствует ризопласт. В жизненном цикле могут образовываться цисты, но в их стенках отсутствует кремний. Цисты прорастают зооспорами. Поселяется как эпифит на нитчатых водорослях. Обитает в стоячих и медленно текущих пресных водоемах.
Класс Рафидовые водоросли – Raphidophyceae
Название класса происходит от греч. rhaphid – игла. Объединяет одноклеточные двужгутиковые организмы, лишенные клеточной стенки. В основном пресноводные организмы с уплощенными клетками (рис. 41).
Клетки имеют Жгутики неравной длины, направленный вперед жгутик – перистый и длиннее, направленный назад – гладкий и короче. Переходная спираль отсутствует. Хлоропласты мелкие, многочисленные или, реже, по 1-2 в клетке, окружены четырьмя мембранами, из них две мембраны ХЭС. Тилакоиды собраны в стопки по три. Ядро крупное, окружено кольцом диктиосом. Митохондрии с трубчатыми кристами. Имеются сократительные вакуоли. Цитоплазма часто бывает вакуолизирована. Клетки голые, окружены только плазмалеммой. Стигмы (глазка) обычно нет.
Фотосинтетические пигменты – хлорофиллы а и с, β -каротин, вошериаксантин, диноксантин, диадиноксантин, гетероксантин.
Запасные продукты ассимиляции – жиры и масла, реже крахмал и хризоламинарин.
Обитают также в сфагновых болотах и других местообитаниях с кислой и нейтральной реакцией воды. Некоторые виды встречаются в солоноватых и морских водах.
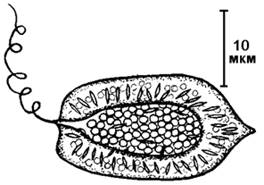
Рис. 41. Внешний вид рафидовой водоросли.
Систематика.
Очень долго эту группу водорослей из-за зеленой окраски, наличия глотки и ряда других особенностей строения рассматривали как класс Хлоромонады среди Евгленовых водорослей. Но данные цитологии, биохимии, физиологии и молекулярной биологии показали их несомненную принадлежность к отделу Гетероконтные (Охрофитовые) водоросли и монофилетичность группы Рафидовых водорослей.
Известно около 25 видов из единственного порядка Хаттонелловые – Chattonellales.
Род Гетеросигма (рис. 42, А) включает морские флагеллаты. Клетки овальной формы со жгутиками, сдвинутыми на боковую сторону, пластиды расположены по периферии клетки. Трихоцист нет. Плавают, совершая вращающие движения вокруг продольной оси тела. Могут вызывать токсичное "цветение воды" в прибрежных водах.
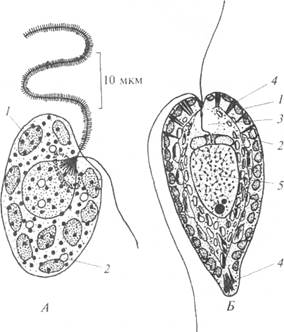
Рис. 42. Рафидовые водоросли (по: R. E. Lee, 1999): А – Гетеросигма; Б – Гониостомум; 1 – хлоропласт; 2 – мукоциста; 3 – сократительная вакуоль; 4 – трихоциста; 5 – ядро
Род Гониостомум (рис. 42, Б) - подвижные дорсовентрально сжатые монады. Спинная сторона выгнутая, брюшная - уплощенная. Борозда на переднем конце ведет в треугольную глотку, из которой выходят два длинных жгутика; их длина сравнима с длиной клетки. Пластиды расположены по периферии клетки. Под плазмалеммой находятся трихоцисты. Обитает в пресных водах с кислотным значением рН.
Класс Евстигмовые водоросли – Eustigmatophyceae
Название класса происходит от греч. eu – «хорошо» и stigma –«метка», «пятно». Класс объединяет голые одноклеточные, реже колониальные, организмы преимущественно коккоидной структуры.
Ядро одно или несколько. Хлоропласт обычно один, окружен 4 мембранами. Жгутик обычно один передний перистый и второе базальное тело или, реже, два неравных жгутика. Стигма (глазок) имеется, расположена за пределами хлоропласта, ее гранулы без мембран.
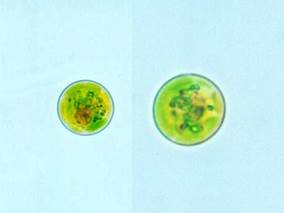
Рис. 43. Внешний вид евстигмовой водоросли
Хлоропласт обычно единственный, крупный, многолопастной, чашеобразный или париетальный, желто-зеленого цвета. Тилакоиды собраны в ламеллы по три, опоясывающая ламелла отсутствует.
Основные пигменты: хлорофилл а, β-каротин, виолаксантин и вошериаксантин. Пиреноид обычно есть только у вегетативных форм.
Запасные вещества – масла и вещество неустановленной природы, которое откладывается в виде твердого материала вне пластид. Крахмал не образуется.
Обитают в пресных водах, реже встречаются в морях и в почве.
Систематика.
Раньше эту группу организмов относили в Желтозеленым водорослям. В настоящее время Евстигмовые водоросли рассматривают в ранге класса в отделе Гетероконтофиты. Известно около 35 видов, преимущественно пресноводных, относимых к единственному порядку Евстигмовые – Eustigmatales.
Род Евстигматос (рис. 43) – одноклеточные мелкие шаровидные водоросли. Клеточная стенка гладкая, цельная, без орнамента. Желто-зеленый хлоропласт один, лопастной, париетальный, со стебельчатыми многогранными пиреноидами. Большая центральная вакуоль с красным содержимым. Размножается 2 или 4 автоспорами или бутылеобразными зооспорами с одним входящим жгутиком, большой стигмой, расположенной за пределами хлоропласта на переднем конце, и единственным задним хлоропластом без пиреноида. Обычный компонент почвенных флор. Известен из Новой Зеландии, Австрии, Исландии, и из бассейнов Аризоны как "горчичная водоросль".
Класс Желтозеленые водоросли – Xantophyceae
К Желтозеленым относятся водоросли, хлоропласты которых окрашены в светло- или темно-желтый цвет, очень редко зеленый и лишь иногда в голубой. Окраска талломов определяется наличием в хлоропластах клеток следующих пигментов – хлорофиллов а и с, β -каротина и ксантофиллов. Преобладание последних и обусловливает своеобразие окраски желтозеленых водорослей. Кроме того, в клетках в качестве основного продукта ассимиляции накапливаются парамилон, капли масла и лишь у некоторых видов, кроме того, глыбки лейкозина и волютина. Крахмал у желтозеленых водорослей не образуется. Отличительной особенностью желтозеленых является наличие у вегетативных клеток монадной структуры и у зооспор двух неравных жгутиков. В состав клеточной стенки входят целлюлоза, глюкоза и уроновые кислоты. Клеточная стенка часто состоит из двух частей.
Размножение вегетативное, бесполое и половое.
Широко распространены в пресных водах. Редко встречаются в морских, солоноватых водах и в почве.
Ранее класс Желтозеленые водоросли назывался Трибофициевые водоросли по типовому роду Трибонема (от греч. tribon – искусный, хитрый и nema – нить). Известно около 450 видов.
Желтозеленые характеризуются значительным морфологическим разнообразием. Среди многочисленных представителей этого отдела обнаруживаются почти все основные типы структуры тела: амебоидная, монадная, пальмеллоидная, коккоидная, нитчатая, разнонитчатая, пластинчатая и сифональная (рис. 44 – 46). Слоевище одноклеточное,
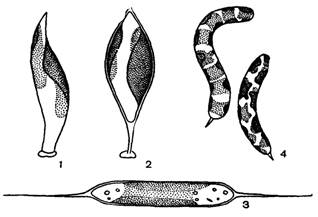
Рис. 44. Внешний вид желтозеленых водорослей: 1, 2 – Харациопсис, 3 – Центритрактус, 4 – Офиоцитиум
колониальное, многоклеточное и неклеточное. Клеточная оболочка плотная, пектиновая и целлюлозная, состоящая из плотно налегающих друг на друга частей или из двух створок. В оболочке откладывается кремнезем или известь. Преимущественно неподвижные формы. Среди одноклеточных видов имеются подвижные формы, лишенные плотной оболочки и снабженные жгутиками, лобоподиями и ризоподиями.
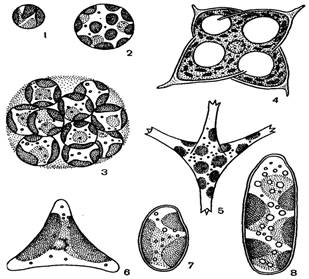
Рис. 45. Внешний вид ксантококковых желтозеленых водорослей: 1–3 – Ботридиопсис, 4 – Тетраэдриелла, 5 – Псевдостаураструм, 6 – Гониохлорис, 7, 8 – Бумиллериопсис
Большинство желтозеленых – неподвижные организмы. У подвижных особей движение может осуществляться с помощью жгутиков либо ризоподий. Клетки различной формы: шаровидные, веретеновидные, эллипсоидные, цилиндрические, тетраэдрические, серповидные, грушевидные, яйцевидные. Талломы размерами от 0.5 – 1.5 мкм (Хлориделла) до нескольких миллиметров в диаметре (Ботридиопсис) (рис. 45, 1 – 3) и до десятков сантиметров в длину (Вошерия) (рис. 46, 3).
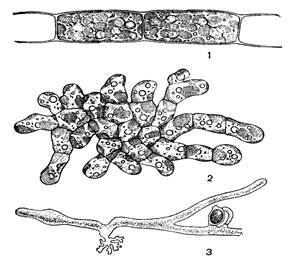
Рис. 46. Внешний вид желтозеленых водорослей: 1 – Трибонема, 2 – Гетеропедия, 3 – Вошерия, часть нити с оогонием и антеридием
Большинство видов желтозеленых являются фототрофами, но встречается и голозойное питание путем заглатывания бактерий и мелких водорослей. Желтозеленые водоросли широко распространены в пресных водах. Они также обычны в почве, реже встречаются в морских и солоноватоводных водоемах. Класс включает аэробионтные, планктонные, донные и перифитонные формы. Эпифиты, эпизоиты, а также внутриклеточные симбионты в клетках простейших.
Независимо от внешней структуры, внутреннее строение клетки желтозеленых водорослей однотипно. В протопласте наблюдается обычно несколько желто-зеленых хлоропластов, имеющих дисковидную, корытовидную, пластинчатую, реже лентовидную, звездчатую или чашевидную форму с цельными или лопастными краями. Окраска связана с отсутствием фукоксантина, отвечающего за золотистый и коричневый цвет у других охрофитовых. Из других пигментов у них имеются β -каротин, вошериаксантин, диатоксантин, диадиноксантин, гетероксантин. У подвижных форм на переднем конце хлоропласта обычно расположен красный глазок, или стигма. У немногих видов имеются пиреноиды полупогруженного типа. Ядро в клетке обычно одно, небольших размеров, но имеются виды и с многоядерными клетками. У некоторых видов в передней части клетки имеется одна или две сократительные (пульсирующие) вакуоли.
У монадных представителей и у подвижных стадий (зооспор и гамет) имеются два неравных жгутика. Исключением являются синзооспоры Вошерии, у которых по поверхности расположены многочисленные пары немного различающихся по длине гладких жгутиков. Короткий жгутик заканчивается акронемой. Жгутики прикрепляются на клетке субапикально. У сперматозоида Вошерии прикрепление латеральное.
У видов с амебоидной, монадной и пальмеллоидной организацией клеточная стенка отсутствует, они покрыты только цитоплазматической мембраной и могут легко менять форму. Иногда «голые» клетки находятся внутри домиков, стенки которых могут быть окрашены в бурый цвет солями марганца и железа. У подавляющего большинства форм имеется клеточная стенка, состоящая из двух частей. В составе клеточной стенки преобладает целлюлоза, а также содержатся полисахариды, состоящие в основном из глюкозы и уроновых кислот. У молодых клеток оболочка тонкая, с возрастом она утолщается. В ней могут откладываться соли железа, соединения которого окрашивают ее в различные оттенки коричневого и красного тонов. Чаще в клеточной стенке присутствует кремнезем, придавая ей твердость и блеск. Она может инкрустироваться также известью и быть различным образом скульптурирована (шипики, ячейки, бородавки, щетинки, зубчики и т.д.) У прикрепленных форм может образовываться вырост оболочки – ножка с прикрепительной подошвой.
У нитчатых форм желтозеленых водорослей с двухстворчатыми оболочками при распаде нитей клеточные оболочки разваливаются на Н-образные фрагменты. Эти фрагменты представляют собой плотно соединенные половинки оболочек двух соседних клеток (рис. 47). При росте нитей Н-образный фрагмент клеточной стенки двух соседних дочерних клеток встраивается между двумя половинками оболочки материнской клетки. В результате каждая из дочерних клеток наполовину покрыта старой оболочкой материнской клетки и наполовину – новообразованной оболочкой.
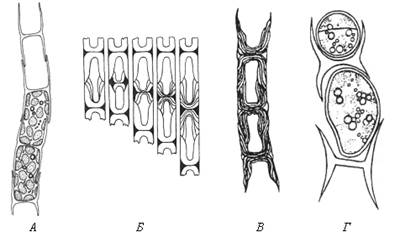
Рис. 47. Схема образования поперечной перегородки между двумя дочерними клетками у нитчатых желтозеленых водорослей (по: А.А. Масюк, 1993): А – фрагмент нити; Б – закладка пояскового кольца и образование поперечной перегородки между двумя клетками; В – слоистость двустворчатых клеточных оболочек; Г – распад оболочки на Н-образные участки
Сократительные вакуоли имеются у подвижных представителей. Обычно их 1-2 на клетку. Аппарат Гольджи своеобразного строения. Диктиосомы мелкие, содержат 3-7 цистерн. Ядро одно, реже их много; у ценотических видов клетки всегда многоядерные.
Размножение. Для большинства видов желтозеленых водорослей характерно вегетативное и бесполое размножение.
Вегетативное размножение осуществляется различными способами: делением клеток пополам, распадом колоний и многоклеточных слоевищ на части. У Вошерии образуются специальные выводковые почки.
При бесполом размножении могут формироваться разнообразные споры: амебоиды, зооспоры, синзооспоры, автоспоры, гемизооспоры, гемиавтоспоры, апланоспоры. Зооспоры «голые» и обычно грушевидной формы.
Половой процесс – изогамия, гетерогамия и оогамия – описан у немногих представителей. У Трибонемы гаметы сходны по размерам, но различаются поведением – это изогамия. У Вошерии наблюдается оогамия: на нитях образуются вместилища женских гамет – оогонии и мужских – антеридии.
В неблагоприятных условиях наблюдается образование цист. Цисты (статоспоры) эндогенные, одноядерные, реже многоядерные. Их стенка часто содержит кремнезем и состоит из двух неравных или, реже, равных частей.
Систематика.
В конце XIX – начале XX вв. различные роды желтозеленых относили к отделу зеленых водорослей, что было связано в первую очередь с окраской и морфологическим сходством талломов. В настоящее время желтозеленые рассматриваются как класс внутри отдела охрофитовых.
Известно около 450 современных видов класса Желтозеленые водоросли, которые группируются в четыре порядка: Ботридиевые, Мишококковые, Трибонемовые и Вошериевые. В основу выделения порядков положены тип дифференциации таллома и особенности жизненного цикла.
Порядок Ботридиевые – Botrydiales. К порядку относятся виды с сифональным типом дифференциации таллома, у которых отсутствует оогамный половой процесс.
Род Ботридиум обитает на почве и имеет вид зеленых пузырей величиной несколько миллиметров, прикрепленных с помощью бесцветных ризоидов. Таллом сифональный, содержит многочисленные ядра и пластиды. Оболочка многослойная, на ней может откладываться известь. Размножение бесполое с помощью двухжгутиковых зооспор, при этом все содержимое пузыря распадается на одноядерные фрагменты. При недостатке влаги размножается с помощью апланоспор или формирует толстостенные цисты. В одних случаях все содержимое пузыря идет на образование одной крупной цисты. В других случаях цисты формируются в ризоидах, куда вначале переходит содержимое пузыря. Цисты прорастают или непосредственно в новый таллом, или образуют зооспоры. Половой процесс – изо- и гетерогамия. Зигота прорастает немедленно, без периода покоя. Обычные и широко распространенные в наземных местообитаниях виды, встречаются по берегам ручьев, прудов или на лишенных растительности почвах.
Порядок Мисхококковые – Mishococcales. Одноклеточные или колониальные представители с коккоидным типом дифференциации таллома.
Род Харациопсис включает одноклеточные прикрепленные формы. При размножении формирует зооспоры, апланоспоры и толстостенные цисты (рис. 44, 1-2).
Род Офиоцитиум (рис. 44, 4) имеет удлиненные цилиндрические клетки, которые могут быть прямыми, согнутыми или спирально закрученными, а на конце могут нести шип. Клеточная стенка состоит из двух неравных частей, из них большая часть участвует в росте клетки, меньшая часть постоянна и имеет форму крышечки. Одноклеточные и колониальные виды, свободноживущие или прикрепленные к субстрату с помощью маленькой ножки. Размножаются зооспорами и апланоспорами, встречаются цисты. Обитают в пресных водах.
Род Мисхококкус образует древовидные прикрепленные колонии. Ветвление дихотомическое и тетрахотомическое. Клетки расположены по 2 или 4 на верхушках слизистых ветвей колонии. Клетки шаровидные, до овальных, с тонкой или толстой клеточной стенкой. Иногда клеточная стенка блестящая и коричневая из-за пропитки ее солями железа. Молодые одноклеточные организмы со слизистым дисковидным основанием, которое служит точкой опоры при прикреплении. После выхода спор протопласт материнской клетки превращается в студень и вытягивается, длина становится в 6 раз больше ширины, и таким образом возникает цилиндрическая ножка. Пустая клеточная стенка материнской клетки всегда становится основанием ножки. Бесполое размножение зооспорами и автоспорами. Автоспоры прикрепляются к верхнему краю слизистой ножки. Последующие клеточные деления повторяют процесс и производят древовидную колонию. Половой процесс – изогамия. Обитают в небольших пресных водоемах как эпифиты нитчатых водорослей. Известны в центральной Европе и Азии.
Порядок Трибонемовые – Tribonematales. Представители имеют нитчатый, разнонитчатый, ложнотканевый и тканевый тип дифференциации таллома. Клеточные стенки либо с Н-образными перекрывающимися частями, либо цельные.
Род Трибонема – неветвящиеся нити (рис. 46, 1). Клетки по форме цилиндрические или бочонкообразные. Клеточная стенка состоит из двух половинок, которые находят краями друг на друга в середине клетки. Оболочки часто слоистые. Фрагменты нитей всегда оканчиваются пустыми половинками Н-образных фрагментов оболочки, имеющих вид вилочки. В клетках несколько желтовато-зеленых пластид, пиреноидов нет. Размножение вегетативное (фрагментацией нитей), бесполое (зооспорами и апланоспорами) и половое (изогамия), причем апланоспоры образуются чаще, чем зооспоры. Могут формировать акинеты. Обитают в пресных водах, где особенно обильно развиваются в холодное время года.
Порядок Вошериевые – Vauсheriales. Все представители имеют сифональный таллом, оогамный половой процесс и синзооспоры.
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 2350; Нарушение авторских прав?; Мы поможем в написании вашей работы!