КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Організм
|
|
|
|
РОЗДІЛ 7
Лекції 11, 12
План
7.1 Клітина (коротке повторення основних положень)
7.2 Від одноклітинних організмів до багатоклітинних
7.3 Від яйцеклітини до багатоклітинного організму
7.4 Диференціювання
7.5 Біологічне старіння
7.6 Гормони
7.1 Клітина (коротке повторення основних положень)
Клітина – структурна й функціональна одиниця будь-якого організму. Гетеротрофні клітини повинні одержувати вуглеводи ззовні, а автотрофні зелені клітини самі створюють їх у хлоропластах шляхом фотосинтезу. Більша частина вуглеводів, що містяться у цитоплазмі, розщеплюється з метою вивільнення енергії – анаеробно (бродіння, ферментація) або частіше аеробно, тобто у процесі окислювання (дихання): спочатку у процесі гліколізу, а потім у циклі лимонної кислоти. Одержувана таким чином енергія зв'язується у формі АТР,більша частина – у ланцюзі дихання, який, так само як і цикл лимонної кислоти, локалізований в мітохондріях. Постачання енергії до автотрофних зелених клітин йде прямим шляхом, без обходу через готові вуглеводи.
Бактеріальні й рослинні клітини самі синтезують усі 20 амінокислот, що входять до складу білків. Тваринним клітинам доводиться одержувати з довкілля принаймні незамінні амінокислоти. Вони поглинають і білки, які розщеплюють потім у лізосомах до амінокислот. Гранулярний ендоплазматичний ретикулум, засіяний рибосомами, синтезує резервні білки й білки, призначені для «експорту», які через систему Гольджі у процесі екзоцитозу залишають клітину. Клітинні й резервні білки зрештою розщеплюються в лізосомах до амінокислот (аутофагія).
Нуклеотиди синтезуються в цитоплазмі. ДНК – носій генетичної інформації – знаходиться в хромосомах, мітохондріях і хлоропластах; там відбувається її реплікація, а також транскрипція, у результаті якої з'являються різні види РНК.
|
|
|
Ліпіди синтезуються в різних компартментах клітини, жирні кислоти – у мітохондріях й у цитоплазмі, а в рослин – головним чином у хлоропластах. Ліпіди мембран також утворюються в мітохондріях і хлоропластах, але головне місце їхнього синтезу – гладкий ендоплазматичний ретикулум. Ліпіди розщеплюються в цитоплазмі (наприклад, у лізосомах); жирні кислоти, що звільнюються при цьому, можуть окислятися в мітохондріях.
7.2 Від одноклітинних організмів до багатоклітинних
Подальший розвиток примітивних одноклітинних привів до трьох різних форм організації: до високодиференційованих одноклітинних (наприклад, найпростіших), до більш великих, багатоядерних, але не розділених на клітини організмів (ценобластів) і до багатоклітинних. До самих високорозвинених життєвих форм прийшли тільки багатоклітинні; цьому сприяла перевага поділу функцій між клітинами з різною спеціалізацією. Перехід до багатоклітинності відбувався у ході еволюції багаторазово і незалежно в різних групах організмів (рис. 7.1, 7.2).
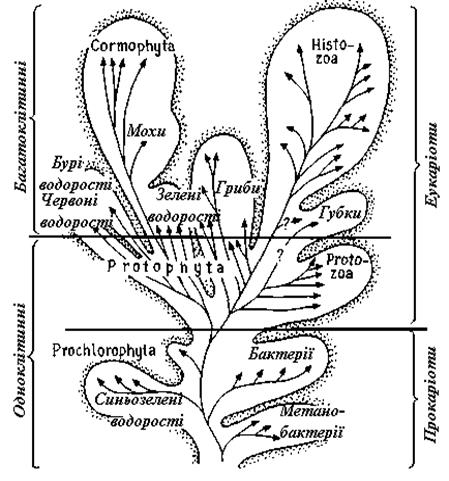
Рис. 7.1. Філогенетичне древо організмів. Поліфілетичне (незалежне в різних групах) виникнення багатоклітинності)
7.2.1 Одноклітинні організми
Одноклітинними є майже усі прокаріоти, протофіти й протозої. Клітина прокаріотичного організму (бактерії й синьо-зелені водорості) не має істинного ядра, мітохондрій, хлоропластів і системи внутрішніх мембран. Їй не властивий мейотичний поділ. Клітинна стінка містить муреїн.
Бактерії бувають кулястими (коки), паличкоподібними (бацили), у формі коми (вібріони) або закрученими гвинтоподібно (спірили) (рис. 7.3). Кулясті бактерії можуть з'єднуватися по двоє (диплококи), у пакети по чотири (сарцини), ланцюжки (стрептококи), грона (стафілококи). Більшість бактерій – гетеротрофи, що харчуються як сапробіонти (мешканці гниючого органічного матеріалу), паразити або симбіонти. Автотрофних бактерій небагато, вони мають здатність до хемосинтезу
|
|
|
ті
| 
|
| А | Б |
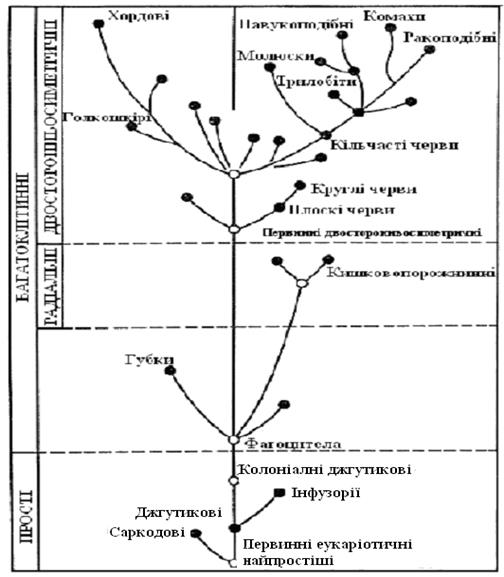
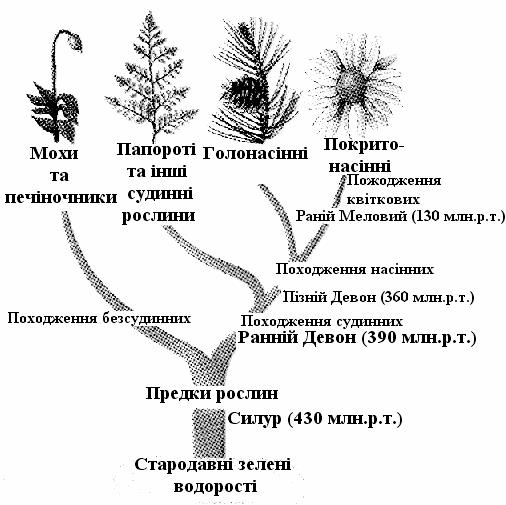
Рис. 7.2. Еволюція еукаріотів. А). Основні етапи еволюції тварин; Б). Філогенетичне дерево рослин
| 
|
| 
|

| 
|
| А | Б |
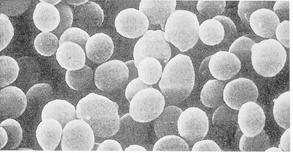
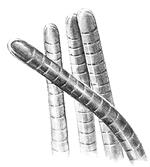
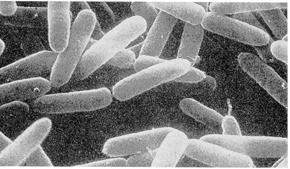
Рис. 7.3. Прокаріотичні одноклітинні організми й об'єднання клітин
А). Основні форми бактерій (електроно-мікроскопічні фотографії):
1 –мікрококи; 2 – паличкоподібні бактерії; 3 – спірили;
Б). Синьо-зелені водорості (х1000)
або (пурпурні й зелені бактерії) до примітивного фотосинтезу без фотосистеми II і, отже, без розщеплення води.
Синьо-зелені водорості (Cyanophyceae) на відміну від бактерій мають «повний» фотосинтез із двома фотосистемами, з фотолітичним розщепленням води й із синіми а іноді червоними фікобіліпротеїдами, але в них ніколи не буває джгутиків.
Архебактерії – древня гілка еволюції первинної прокаріотичної клітини. Одні з архебактерій – метанові – одержують енергію за допомогою реакцій, при яких виділяється метан, наприклад СО2 + 4Н2 → СН4+2Н2О або СН3СООН → СН4+СО2. Від усіх інших живих організмів вони відрізняються, крім іншого, незвичайними коферментами, незвичайною структурою t PHK й r РНК, клітинною стінкою без характерного для прокаріот муреїну. Процеси, які здійснюють метанобактерії, використовують для переробки органічних відходів в анаеробних умовах з одержанням енергоносія. Метанобактеріям дуже близькі галофільні бактерії солоних озер і кислото- і термофільні бактерії, що окислюють сірку, з кислих гарячих джерел.
Протофіти (одноклітинні рослини) – це не таксономічна група, а рівень організації, представники якої є серед усіх груп водоростей (крім бурих) і грибів. Клітини протофітів (еукаріотичні) не мають клітинної стінки або ж мають стінку із целюлози (типову для рослинних клітин), або з хітину. Вони гетеротрофні (гриби) або автотрофні (водорості); в останньому випадку вони мають один або кілька хлоропластів.
Протозої (найпростіші – одноклітинні тварини) гетеротрофні. Корененіжки (Rhizopoda, рис. 7.4)використовують для пересування й захоплення здобичі жертви
|
|
|
виступи (псевдоподії), що утворюються тимчасово. Джгутикові (Zooflagellata) не мають хлорофілу; пересуваються за допомогою 1, 2, 4 або більшого числа джгутиків. Протозої використовуються як індикаторні організми для оцінки якості води в природних водоймах і для оцінки ефективності роботи активного мулу в біологічних очисних спорудах. Присутність джгутикових найпростіших характеризує воду як «сильно брудну», амеб – як «брудну», вільно плаваючих інфузорій – як «відносно чисту», прикріплених інфузорій – як «чисту».
7.2.2 Ценобластична організація
Деякі водорості й гриби мають багато ядер, але не розділені на клітини. В їхніх вегетативних тілах, що найчастіше мають трубчасту форму та можуть бути досить великі й високоорганізовані (рис. 7.4),кожне ядро утворює разом зі своєю «сферою впливу» у протоплазмі енергіду.

| 
| 
| 
|
| 
| 
| 
|
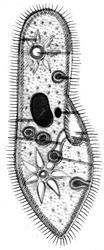
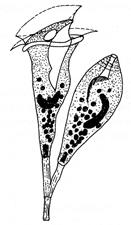
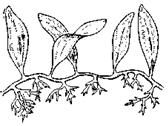
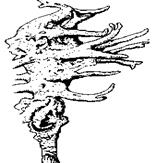
Рис. 7.4. Одноклітинні організми й об'єднання клітин
Найпростіші: 1 – Корененіжки (Amoeba, х200); 2 – Джгутикові (Bodo saltans, х200); 3 –Вільноплаваючі інфузорії (Paramecium, х200); 4 – Прикріплені інфузорії (Epistylis, х200). Ценобласт: 5 – Caulerpa (х1/3) Ценобії; 6 – Dinobryon (х250); Агрегаційні об'єднання: 7 – Scenedesmus (х600); Колонії: 8 – Volvox (х400)
Плазмодії – багатоядерні маси протоплазми, що утворюються як результат злиття голих клітин (істинні плазмодії у слизових грибів – міксоміцетів) або просто їхнього угруповання (псевдоплазмодії у міксобактерій). Вони можуть як єдине ціле повільно перетікати або переповзати з місця на місце.
7.2.3 Об'єднання клітин
Об'єднання клітин являють собою проміжні кроки на шляху до багатоклітинних організмів.
Ценобії – комплекси слабко об'єднаних клітин – виникають у результаті клітинного поділу. Клітини залишаються зв'язаними матеріалом, подібним до студену (рис. 7.4). Агрегаційні об'єднання утворюються як результат тимчасового з'єднання одноклітинних організмів. Вони можуть як завгодно розпадатися на більш дрібні одиниці. Колонії виникають шляхом клітинного поділу. Клітини в них утворюють морфологічну й функціональну єдність (рис. 7.4). Їхні цитоплазми з'єднані між собою й можуть координовано реагувати (синхронне биття джгутиків). У високоорганізованих колоніях, наприклад у кулястій водорості Volvox (рис. 7.4), спостерігається розподіл функцій (є вегетативні клітини, що забезпечують рух, і генеративні, які потрібні для розмноження), отже їх можна вважати першими багатоклітинними організмами.
|
|
|
7.2.4 Багатоклітинні організми без справжніх тканин
Усі багатоклітинні рослини можна об'єднати під назвою Metaphyta (сюди входять Thallophyta, Bryophyta й Cormophyta), а тварин – під назвою Metazoa (Porifera й Histozoa). Bryophyta, Cormophyta й Histozoa мають справжні тканини.
Талофіти (Thallophyta). Талом – це багатоклітинне (або ценобластичне) вегетативне тіло, що не розділене на корінь, стебло і листя. Талофіти – рослини з таломом – не складають єдиної таксономічної групи, а входять до складу різних груп водоростей і грибів (рис. 7.5). У нитчастих таломах клітини, розмножуючись поперечним поділом, утворюють один ряд. Плоскі таломи виникають із нитчастих в результаті поздовжнього поділу клітин. У більш високорозвинених форм з'являється полярна диференціація талома, розгалуження талома. Талом грибів – міцелій – складається найчастіше з безлічі ниток – гіф, які рясно розгалужуються.
| 
| 
| 
|
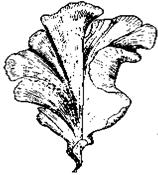
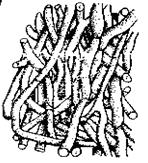
Рис. 7.5. Талофіти й губки
1 – зелена водорість Ulva (х1/3); 2 – плектенхіма в ніжці плодового тіла білого гриба, Boletus edulis (х200); 3 – перетин поперечний зріз лишайника
Collema (х200); 4 – прісноводна губка Spongilla lacustris (хI/2)
Таломні нитки можуть поєднуватися у псевдотканини (у червоних водоростей, у плодових тілах вищих грибів): переплітаючись, вони утворять плектенхіму (плетену тканину), а зростаючись – псевдопаренхіму.
Лишайники – це симбіотичні автотрофні об'єднання водорості (синьо-зеленої або зеленої) із грибом (рис. 7.5).
Губки (Porifera) крім клітин, що служать для розмноження, мають різні типи соматичних клітин, які, однак, не поєднуються в органи або справжні тканини (рис. 7.5). Тільки на поверхні тіла клітини можуть утворювати подобу епітелію. Але їхнє скріплення пухке, і вони легко можуть вийти з об'єднання. Більшість клітин здатна до амебоїдному руху. Губки – прикріплені (сидячі), в основному морські організми.
7.2.5 Багатоклітинні організми із справжніми тканинами
Тканини – це об'єднання клітин, подібних за будовою й функцією.
Кормус – це вегетативне тіло, яке складається із кореню й пагону (пагін = стебло + листя). До кормофітів, рослин з кормусом, відносяться папоротеподібні (Pteridophyta) й насінні рослини (Spermatophyta). Bryophyta (мохи) займають проміжне положення між талофітами й кормофітами. Їхнє вегетативне тіло або являє собою талом, або розділено на стебло і листя, але завжди має замість кореня ниткові одноклітинні ризоїди.
Утворення тканини починається з верхівки талома, пагона або кореня (коли останній є). Тканини, що забезпечують утворення (меристеми) залишаються ембріональними тканинами, здатними до росту за рахунок поділу клітин. Верхівкові меристеми перебувають на кінчиках пагонів і коріння, а вторинні і залишкові меристеми – усередині пагонів і коріння.
Постійні тканини (рис. 7.6) – зрілі тканини. До них відносяться порівняно мало спеціалізовані основні тканини (паренхіма, що служить, наприклад, для фотосинтезу або накопичення запасних речовин), покривні тканини (наприклад, епідерміс і коркова тканина, що служать для ізоляції від зовнішнього середовища), тканини, що всмоктують (наприклад, епідерміс кореня), необхідні для поглинання яких-небудь речовин, тканини, що проводять (наприклад, ситовидні трубки для транспорту продуктів асиміляції, судини для транспорту води), механічні тканини (наприклад, склеренхіма, що має опорну функцію), видільні тканини (наприклад, залози) і репродуктивні тканини (спороутворюючі й пилокоутворюючі).
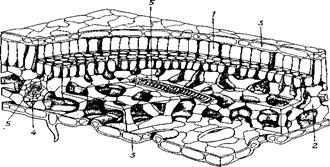
Рис. 7.6. Тканини у Cormophyta: анатомія листа
1 – палісадна паренхіма (фотосинтезуюча тканина); 2– губчата паренхіма; 3 – епідерміс; 4 – ситовидні трубки; 5 – судини
Histozoa – багатоклітинні тварини, клітини яких, поєднуючись, утворюють тканини, хоча б покривні (епітелії). Всі ці тварини, за винятком кишковопорожнинних (Соеlenterata), мають органи.
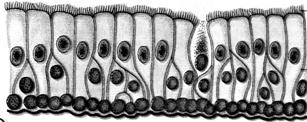
Покривні, або епітеліальні, тканини - це шари щільно покладених клітин, що покривають поверхню тіла або порожнини (рис. 7.7). Відмерлі або відторгнуті епітеліальні клітини постійно заміщаються в результаті проліферації клітин, що зберігають здатність ділитися (фізіологічна регенерація). Від епітеліальних клітин походять залозисті клітини (залозисті тканини).
Сполучні тканини – це пухкі об’єднання клітин, що заповнюють проміжки між органами. Клітини виділяють міжклітинну (основну) речовину. У сполучній тканині вона м'яка й може містити колагенові або еластичні волокна, розташовані безладно, паралельно один одному (у сухожиллях) або хрест-навхрест (у фасціях).
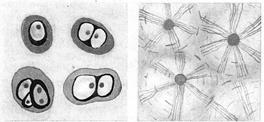
Для опорних тканин характерна міцна й щільна міжклітинна речовина. У хрящовій тканині вона еластична при натисненні, гнучка, у ній немає кровоносних судин. У кістковій тканині (властивої хребетним) через відкладення солей кальцію міжклітинна речовина набуває твердість і містить гаверсові канали із кровоносними судинами й нервами. Кісткові клітини (остеоцити) розташовуються в основному концентричними рядами навколо гаверсових каналів і зв'язані між собою плазматичними відростками (рис. 7.7).
| 
| |||

| 
| |||
Рис. 7.7. Тканини у Histozoa.
1 – війчастийий епітелій; 2 – хрящова тканина; 3 – кісткова тканина; 4 – гладка м'язова тканина; 5 – поперечносмугаста м'язова тканина; 6 – уніполярний нейрон; 7 – біполярний нейрон;8 – мультиполярний нейрон
М'язова тканина складається з клітин з дуже сильно розвинутою здатністю до оборотного скорочення. Гладкі м'язові клітини здебільшого мають веретеновидну форму, містять одне або багато ядер, довжина цих клітин до 0,5 мм. Поперечносмугасті м'язові клітини ( м'язові волокна ) мають довжину до 12 см і містять багато ядер. Деякі безхребетні мають м'язові гвинтоподібні фібрили в косопокресленої мускулатурі.
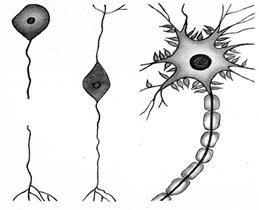
Клітини нервової тканини (нейрони) сприймають, зберігають і переробляють інформацію. Тіло нейрона (перикаріон)має один, два або більше число відростків (уні-, бі- і мультиполярні нервові клітини, рис. 7.7). В останньому випадку, як правило, короткі, товсті, сильно розгалужені відростки (дендрити) проводять збудження до перикаріону, а один дуже довгий відросток ( нервове волокно, нейрит,або аксон) – від перикаріону.
Розрізняють немієлінізовані, слабко мієлінізовані й мієлінізовані нервові волокна. В останніх мієлінова оболонка – шар, який містить ліпіди з 100-200 обмотаних навколо аксона мембран з високим електричним опором – через кожні 1-3 мм переривається (перехоплення Ранв’є). Його утворюють швановскі клітини (рис. 7.8). Аксони можуть об’єднуватися в "тонкі" пучки, а потім – у більш товсті пучки, утворюючи нерв. Аферентні (чутливі, центрипетальні) нерви проводять збудження від периферії до центру, а еферентні (ефекторні, центрифугальні) – у зворотному напрямку. Змішані нерви містять як аферентні, так і еферентні волокна.
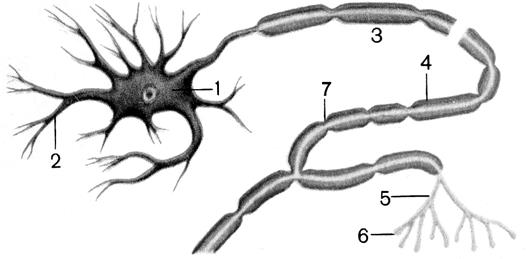
Рис. 7.8. Будова нейрона
1 – тіло нейрона; 2 – дендрити; 3 – аксон; 4 – мієлінова оболонка; 5 – нервові закінчення аксона; 6 – синоптична бляшка; 7 – перехоплення Ранвьє
На розгалужених кінцях аксонів знаходяться кінцеві ґудзички (рис. 7.8); це місця контактів (синапси) для передачі збудження на наступний нейрон або іншу клітину (наприклад, залозисту або м'язову). У синапсі один проти одного розташовані ділянка передсинаптичної мембрани і постсинаптична мембрана приймаючої клітини. Між ними перебуває синаптична щілина (20-35 нм).
7.2.6 Структурна й функціональна організація вищих організмів
Вищі рослини (кормофіти) в основному автотрофні. Внаслідок цього їм необхідні великі поверхні, що поглинають світло (листя, розгалуження пагонів) (рис. 7.9). Листя розташоване таким чином, що майже не затінюють один одного; згинання пагона в напрямку до світла (фототропізм) і розташування палісадних клітин на зразок «світлової шахти» (рис. 7.6) збільшують поглинання світла. У кормофітів переважає радіальна симетрія.
У наземних рослин розвиток зовнішньої поверхні значно більше, ніж у тварин, що приводить до величезних втрат води в результаті транспірації. Ці втрати повинні бути доповнені процесом поглинання води з ґрунту. З кореня через стебло йдуть у листя провідні пучки. Частина пучка, звичайно внутрішня (ксилема, або деревина), містить судини (мертву провідну тканину) для транспорту води (рис. 7.6, 7.9). Властивість сухого повітря – це дуже низький водний потенціал, а ґрунту – високий. Вода завжди рухається убік більше низького водного потенціалу. Сила смоктання, обумовлена транспірацією, створює в судинах негативний тиск, і стовп води рухається нагору. Розриву водяного стовпа в стовбурах високих дерев перешкоджає сила зчеплення між молекулами води. Рослина одержує вуглець (у вигляді СО2) з повітря, а інші живильні речовини у вигляді іонів – із ґрунту.
Інша частина провідного пучка, звичайно зовнішня – флоема, або луб (рис. 7.6, 7.9) – містить ситовидні трубки (провідну тканину) для транспорту асимілятів, що утворюються в процесі фотосинтезу. Виникаючий надлишок тиску (потенціал тиску) жене розчин асимілятів (відповідно до теорії течії рідин) до місць їхньої витрати й накопичення (до осі пагона, кореню, квіткам, плодам). Там відбувається активне поглинання асимілятів і завдяки цьому підтримуються градієнти концентрації, градієнти осмотичного потенціалу й потенціалу тиску.
Для потужного розгалуження пагона необхідна висока міцність його осі. Живі (коленхіма) і мертві (склеренхіма) механічні тканини з їх товстими, часто здерев'янілими клітинними стінками зрідка розташовуються у формі суцільних або переривчастих кілець у зовнішніх частинах (корі) стебла (за принципом сталевої труби). Вторинне зростання у товщинувідбувається за рахунок камбію – внутрішньої меристеми. Камбій знаходиться між флоемою та ксилемою у вигляді суцільного кільця й утворює із зовнішньої сторони флоему з ситоподібними трубками, а із внутрішньої – ксилему з судинами (рис. 7.9, Б).
Реагуючи на фактори зовнішнього середовища, наприклад на тривалість щоденного світлого періоду (існують рослини короткого й довгого світлового дня) або на температуру, рослина, що вегетує, переходить у репродуктивний стан: у меристемі на кінцях пагонів зав’язуються квітки, а після запилення й запліднення розвиваються насіння й плоди.
На відміну від багатоклітинних рослин багатоклітинні тварини (Metazoa), за винятком губок, мабуть, виникли від одноклітинних предків тільки одним шляхом. Серед тварин великого різноманіття та високого розвитку досягли тільки тварини з ціломом – вторинною порожниною тіла, до яких відносяться всі типи тварин (рис. 7.10, 7.11), починаючи з пласких черв’яків.
| 
|
| Б | |

| |
| А | В |
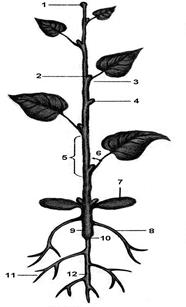
Рис. 7.9. Розвиток насінної рослини
A. Будова молодої дводольної рослини: 1 – верхівкова брунька, 2 – пазушна брунька, 3 – вузол, 4 – придаткова брунька, 5 – міжвузля, 6 – пазуха листа, 7 – сім'ядоля, 8 – придатковий корінь, 9 – підсім'ядольне коліно, 10 – коренева шийка, 11 – бічні корені, 12 – головний корінь. Б. Зріз молодого пагона: 1 – пучковий камбій, 2 – міжпучковий камбій, 3 – вторинна ксилема, 4 – вторинна флоема. В. Зріз старого пагона: 5 – первинна флоема, 6 – вторинна флоема,7 – васкулярний камбій, 8 – вторинна ксилема, 9 – первинна ксилема, 10 – серцевина, 11 – первинний серцеподібний промінь, 12 – вторинний серцеподібний промінь
Тварини ведуть рухливий, рідше прикріплений (тільки водні форми) або паразитичний спосіб життя. Щоб вільно пересуватися, їм, на відміну від рослин, необхідні мінімальна поверхня та компактна форма тіла. Рано з'являються білатеральна симетрія тіла і його витягнутість у напрямку руху. Міцність забезпечують кістякові утворення, які перебувають або на поверхні тіла (екзокістяк) – у молюсків, членистоногих та ін., або усередині (ендокістяк) – у голкошкірих, хордових та ін. У прикріплених форм, таких як кишковопорожнинних, більш звичайна радіальна симетрія.
|
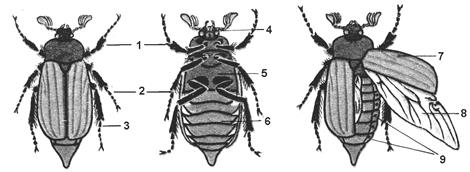
Рис. 7.10. Хрущ. Зовнішня будова (схема): 1 – перша пара ніг, 2 – друга пара ніг, 3 – третя пара ніг, 4 – голова, 5 – груди, 6 – черевце, 7 – надкрила, 8 – крило, 9 – дихальця;
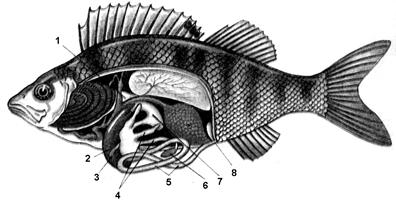
Рис. 7.11. Травна система окуня
1 – стравохід, 2 – печінка, 3 – жовчний міхур, 4 – придатки кишечнику,
5 – кишечник, 6 – селезінка, 7 – шлунок, 8 – анальний отвір
Всі тварини – гетеротрофи, тобто у своєму харчуванні прямо або побічно залежать від первинних продуцентів. Серед них розрізняють фітофагів (рослиноїдних), зоофагів (хижаків) і сапрофагів (що харчуються трупами організмів). Харчуючись, тварина одержує джерела енергії (вуглеводи, жири, білки) і речовини, важливі у функціональному відношенні (вітаміни, деякі ліпіди, мінеральні речовини, воду
Поглинені енергетичні речовини в процесі травлення розщеплюються на порівняно низькомолекулярні фрагменти, які можуть всмоктуватися. Цей процес у вищих тварин відбувається винятково або частково в шлунково-кишковому тракті ( позаклітинне травлення) за участю травних ферментів, які утворюються в самому кишковому епітелії та у великих придаткових залозах (у слинних і підшлункових залозах у хребетних, у залозах середньої кишки у безхребетних). Після попереднього позаклітинного переварювання провадиться більш повне внутрішньоклітинне. Усмоктування низькомолекулярних продуктів травлення, а також функціонально важливих речовин із просвіту кишечнику в тіло відбувається пасивним та активним способом (у хребетних головним чином у тонкому кишечнику). Неперетравлені, непридатні для засвоєння речовини викидаються при дефекації у вигляді калу (фекалій).
Транспорт речовин, що всмокталися, здійснюється головним чином шляхом конвекції в системі трубок, якими циркулює рідина (кров, гемолімфа) – у кровоносній системі. У примітивних системах стінками судин проходять перистальтичні хвилі, що проштовхують кров уперед. У більш високорозвинених організмів відокремлюються певні пульсуючі ділянки судин – серця. Серце працює ритмічно, фаза скорочення (систола) регулярно переміняється фазою розслаблення, або наповнення (діастолою). Циркуляція відбувається або винятково в судинах ( замкнута система кровообігу ), або в деяких ділянках кров виходить із судин і тече по лакунах між тканинами (незамкнута система кровообігу),як у членистоногих, молюсків та ін.
Для аеробного розщеплення живильних речовин з метою одержання енергії необхідний кисень. Дуже дрібним тваринам (наприклад, плоским чирвам, нематодам, коловерткам, кліщам) для життя досить кисню, що надходить через поверхню тіла (шкірне дихання). Але чим крупніше тварина, тим не вигіднішим стає відношення поверхні тіла до його об’єму. З'являється необхідність в органах дихання. Зябра – це тонкостінні листоподібні, нитковидні або пір'ясті вирости поверхні тіла, у яких в основному й відбувається газообмін. У наземних тварин зябра з їхнім ніжним покривом легко висихали б. Тому органи повітряного дихання являють собою вгинання. Легені – це тонкостінні порожнини, у які вільно проникає повітря. Легені захищені стінками грудної порожнини. Вони забезпечуються свіжим повітрям шляхом вентиляції (вдихів і видихів) через ротове та (або) носові отвори. Трахеї є другою основною формою органів повітряного дихання, яка властива багатьом членистоногим. Це розгалужені повітряні канали, які відкриваються на поверхні тіла отворами (дихальцями), і підходять безпосередньо до дихаючих тканин.
Для видалення отрутного NH3, продукту розпаду білків, потрібні особливі механізми. Тільки водні тварини віддають NH3 у незміненому вигляді в навколишнє середовище.Наземні тварини, витрачаючи енергію, зв'язують NH3 у неотруйні сполуки. Наприклад, амфібії й ссавці синтезують із NH3 головним чином сечовину,а комахи, рептилії й птахи – сечову кислоту.
Органами виділення можуть служити: протонефридії, нефридії, мальпігієві судини. У хребетних для більш ефективної фільтрації виникли щільні сплетіння кровоносних судин з канальцями видільної системи – мальпігієві клубочки.У результаті фільтрації спочатку утворюється первинна сеча, з якої під час її проходженні через канальці (сечовики) шляхом зворотного усмоктування (реабсорбції) витягуються потрібні речовини, а інші речовини вводяться в неї шляхом секреції, у результаті виходить дефінітивна сеча.
Для координації й керування функціями у вищих тварин служать дві комунікаційні системи– нервова й гормональна. Найпростішу, дифузійну нервову системумають кишковопорожнинні: під ектодермою у них залягає плоска мережа з мультіполярних нейронів, що можуть проводити збудження у всіх напрямках. У тваринному світі рано з'являються місцеві скупчення нервових клітин, нервова система підрозділяється на центральну.При цьому одна еволюційна лінія веде до вентрального (черевного) нервового ланланцюжка (тварини з вентральною нервовою системою), а інша – до дорсальної нервової трубки, що утворюється у разі вгинання ектодерми (тварини зі спинним мозком). У тварин з білатеральною симетрією на передньому кінці тіла, де розвиваються великі органи чуття, відбувається значна концентрація нервової тканини (цефалізація).Центральні функції керування бере на себе головний мозок.
Завдяки нервовим клітинам, які, подібно «датчикам» у системах регуляції, стежать за параметрами внутрішнього середовища, тваринний організм може за допомогою тонких регуляторних механізмів підтримувати сталість внутрішнього середовища. Найвищого розвитку ці системи гомеостазу досягають у птахів і ссавців. Підтримуючи постійну порівняно високу температуру тіла (гомойотермність), ці тварини створюють незалежні від зовнішнього середовища внутрішні умови, сприятливі для роботи нервової системи та інших органів.
Тварини, особливо високоорганізовані (хребетні, членистоногі, головоногі молюски), найчастіше роздільностатеві; самець має сім’яники, самка – яєчники, у яких дозрівають статеві клітини. Гермафродити зустрічаються особливо часто серед пласких черв’яків, олігохет, легеневих брюхоногих молюсків та оболочників. Самці й самки нерідко розрізняються за будовою й розмірами тіла (статевий диморфізм).
7.3 Від яйцеклітини до багатоклітинного організму
Кожний багатоклітинний організм розвивається у результаті безлічі мітозів з одноклітинної стадії – заплідненної яйцеклітини
7.3.1 Розвиток багатоклітинної рослини
У насінних рослин запліднена яйцеклітина (зигота) розвивається в насінний зачаток (рис. 7.16). Із зиготи в результаті поперечних поділів утворюється нитковидний передзародок. Його верхівкова клітина дає початок зародку, а інші перетворюються в підвісок. Після ряду поздовжніх і поперечних поділів верхівкової клітини виникає восьмиклітинне кулясте утворення (рис. 7.16). У результаті тангенціальних поділів утворюється 8 внутрішніх і 8 зовнішніх клітин; із зовнішніх клітин пізніше розвиваються покриви (епідерміс пагона, епідерміс кореня). З верхівкових частин ембріона формуються зародкові листя (сім'ядолі) і маленька брунечка (плюмула – верхівкова меристема стеблинки), а з базальних частин – підсім’ядольне коліно, або гипокотиль, корінець (радикула) і його захисна шапочка – кореневий чохлик (каліптра) (рис. 7.12).
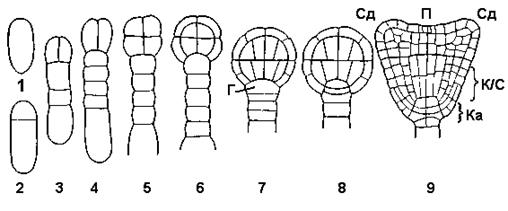
Рис. 7.12. Розвиток зародку у пастушій сумці
Сд – сім'ядолі; П – брунечка; К/С – корінець та стеблинка; Ка–каліптра (кореневий чохлик); Г – гіпофіза (верхня клітина підвіска)
Під час проростання насіння з його оболонки насамперед проривається корінець із чохликом, а потім уже надземні частини. Сім'ядолі незабаром зникають, а із плюмули виникає вся система пагонів.
7.3.2 Розвиток багатоклітинної тварини
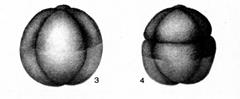
Ембріональний розвиток охоплює проміжок часу від першого поділу яйця до вилуплення або народження молодої особини. У результаті мітотичних поділів заплідненої яйцеклітини утворюються бластомери. У бідних жовтком яєць ділиться вся маса яйця (повне дроблення). Перша площина дроблення проходить, як правило, меридіонально, тобто уздовж осі яйця, друга – перпендикулярно до першого й теж меридіонально, а третя екваторіально (рис. 7.13).
| |
| 
|
Рис. 7.13. Дроблення яйцеклітини
1 – починає формуватися перетяжка між двома першими клітинами, на які ділиться яйцеклітина, 2 – сформувалося два перших бластоміри, 3 – чотири бластоміри, 4 – вісім бластомірів
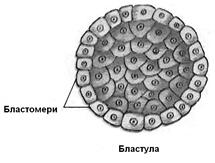
У результаті дроблення утворюється бластула. У типовому випадку це наповнена рідиною порожня кулька (целобластула) з епітелеподібною стінкою (бластодермою) із бластомірів і центральною порожниною (бластоцелем – первинною порожниною тіла).
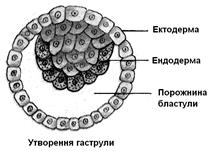
Із бластули утвориться гаструла – чашоподібний зародок із двох шарів (зародкових листків), ектодерми (зовнішній шар) і ендодерми (внутрішній). У найпростішому випадку гаструляція відбувається шляхом вгинання бластодерми (інвагінація, рис. 7.14), причому отвір, що залишається після вгинання, стає первинним ротом, а внутрішня порожнина – первинною кишкою.
|
Рис. 7.14. Гаструляція
А. Вгинання (інвагінація); 1 – порожнина бластули, 2, – ектодерма, 3, – ендодерма, 4 – первинний рот, 5 – порожнина первинної кишки
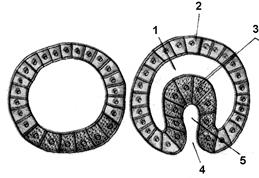
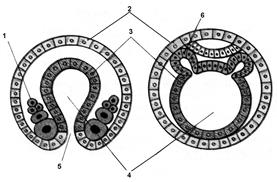
Губки й кишковопорожні зупиняються на стадії двох зародкових листків. В інших багатоклітинних тварин утвориться ще третій зародковий листок – мезодерма. Його закладка формується у вигляді епітеліального шару з ентодерми та розташовується між екто- і ентодермою. У багатьох типів тварин мезодерма виникає шляхом відщеплення від «даху первинної кишки». Порожнина, вистелена мезодермою, – це, так називана, вторинна порожнина тіла (целом) (рис. 7.15).
| 
|
| А | Б |
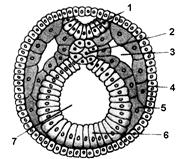
Рис. 7.15. Розвиток ембріона хребетних
А. Тілобластичний тип утворення мезодерми: 1 – мезодерма, 2 – ектодерма, 3 – ендодерма, 4 – порожнина первинної кишки, 5 – первинний рот, 6 – мезодерма. Б. Нейрула. Гисто- і органогенез: 1 – нервова пластинка, 2 – хорда, 3 – вторинна порожнина тіла, 4 – ектодерма, 5 – мезодерма, 6 – ендодерма, 7 – порожнина кишки
Під час подальшого розвитку ембріона та диференціації з ектодерми утворюються: покривна тканина (епідерміс) з її залозами й похідними структурами (кутикула, хітин, вапняні раковини, волосся, пір'я, луска, пазурі, нігті, копита й т.п.), сенсорні епітелії органів почуття, кришталик ока, нервова система, у тому числі задня частка гіпофіза.
З мезодерми утворюються: стінка цілома (у хребетних – очеревина, плевра й брижі), основна частина мускулатури, сполучна й опорна тканина, кровоносні судини, у тому числі серце, клітини крові й лімфи, нефридії, нирки хребетних тварин, сім'япроводи і яйцепроводи.
З ентодерми утворюються: епітелій середньої кишки з його придатковими залозами (печінка й підшлункова залоза в хребетних, залози середньої кишки в багатьох безхребетних), у хордових – хорда, зяброві кишені і їхні похідні (легені, плавальний міхур, тимус та ін.), а також щитовидна залоза.
Основні стадії ембріогенезу людини представлені в табл. 7.1.
Таблиця 7.1
|
|
|
|
|
Дата добавления: 2015-05-24; Просмотров: 2481; Нарушение авторских прав?; Мы поможем в написании вашей работы!