КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Юрский экологический кризис сообщества наземных тетрапод
|
|
|
|
Юрский экологический кризис сообщества наземных тетрапод и эвристическая модель сопряженной эволюции сообщества и биоты
Проблемы доантропогенной эволюции биосферы. Москва: "Наука", 1993. С. 60-95.
Эта статья подготовлена в соответствии с программой «Проблемы эволюции ведущих компонентов биосферы» (Соколов, 1988). В первой части, посвященной юрскому кризису, она продолжает исследования по филоценогенезу, проводившиеся в лаборатории членистоногих Палеонтологического института РАН (Жерихин, Сукачева, 1973; Калугина, 1974, 1977; Калугина, Жерихин, 1975; Жерихин, Калугина, 1980, 1985; Пономаренко, Калугина, 1980; Назаров, 1984; Пономаренко, 1984, 1986; Дмитриев, Жерихин, 1988), но на материале наземных (неморских) тетрапод. Во второй части статьи предложена модель, развивающая представления В. В. Жерихина о ценотической регуляции филогенеза (Родендорф, Жерихин, 1974; Жерихин, 1978—1987; Жерихин, Расницын, 1980; Расницын, 1988— 1990). Она, в частности, помогает понять природу крупных экологических кризисов, подобных юрскому.
Результаты наших исследований по филоценогенезу отражены в ряде предварительных публикаций (Каландадзе, Раутиан, 1979— 1984; Раутиан, 1980; Раутиан, Каландадзе, 1989). В развитие комплекса работ палеоэнтомологов и наших, но на рецентном материале вышла серия публикаций А. А. Вахрушева (1984—1988).
Обнаружение юрского кризиса связано с нашими исследованиями по исторической зоогеографии (Каландадзе, 1974; Каландадзе, Раутиан, 1977—1990; Раутиан, Каландадзе, 1984). Результатом их, в частности, явилась реконструкция зоогеографических областей прошлого для наземных (неморских и нелетающих) тетрапод (рис. 1), которая впервые дает последовательное палеонтологическое обоснование современным зоогеографическим областям А. Р. Уоллеса (Элтон, I960; Wallace, 1876). Наши реконструкции произведены по эпохам, начиная с ранней перми и кончая плейстоценом, исключая лишь раннюю и среднюю юру. Последнее связано с феноменальной бедностью фаун этого времени и настолько слабыми фаунистическими связями, что мы не рискнули дать зоогеографическую реконструкцию на столь скудном материале. При этом географическая представленность местонахождений наземных тетрапод оказалась удивительно широкой: кроме Антарктиды, они представлены на всех материках, включая Мадагаскар (Каландадзе, Раутиан, 1983). Сходная ситуация наблюдалась лишь на заре экологической экспансии тетрапод в девоне, когда их остатки известны из Северной и Южной Америки, Гренландии, Восточной Европы, Сибири и Австралии, но их фауна еще беднее и состоит из уникальных для каждого местонахождения форм (Лебедев, 1985). Пример климатического обеднения дает фауна карбона, когда появляется выраженная климатическая зональность, отсутствовавшая в девоне (Вахрамеев и др., 1970; Мейен, 1984, 1988). Фауна тетрапод карбона представлена только в Северной Америке и Западной Европе, что, очевидно, связано с расположением этих стран в тропической зоне (Вахрамеев и др., 1970; Мейен, 1984, 1988), нахождением Северного полюса на северо-востоке современной Сибири и оледенением Гондваны в позднем карбоне (Вегенер, 1984; Рид, Уотсон, 1981). Она значительно богаче ранне-среднеюрской и в пределах фаунистически охарактеризованных стран вполне поддается зоогеографической интерпретации (Каландадзе, Раутиан, 1980, 1981). Уникальность юрского события для всего интервала, начиная с карбона и до современности, заставила проанализировать его более подробно.
|
|
|
Рис. 1. Реконструкция фаунистических (историко-зоогеографических) областей прошлого по наземным тетраподам
Динамика таксономического разнообразия семейств и родов наземных тетрапод (рис. 2) обнаружила для ранней—средней юры абсолютный минимум разнообразия для всей их палеонтологически документированной истории начиная с позднего карбона. Разнообразие тетрапод в это время оказалось сопоставимым с таковым для среднего карбона Северной Америки и Западной Европы.
|
|
|
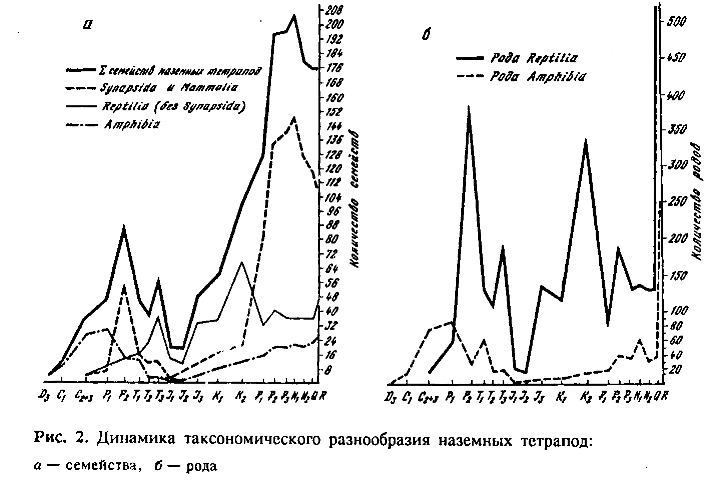
Мы также проанализировали динамику впервые и в последний раз встречающихся в палеонтологической летописи таксонов. Оказалось, что при грубом рассмотрении (с точностью до эпохи и не ниже родового уровня) динамика появляющихся и исчезающих таксонов не обнаружила качественных отличий от таковой для существующих в каждый данный момент времени семейств и родов. Этот результат хорошо согласуется с укоренившимся представлением об относительно высоком среднем темпе эволюции тетрапод и быстрой смене их таксономического разнообразия во времени (Симпсон, 1948; Расницын, 1987). Количество появляющихся и исчезающих семейств и родов в первом приближении пропорционально наличному в данный момент разнообразию. По этой же причине на нашем материале не дали эффектных результатов расчеты разного рода индексов когерентности и оборота фауны, с успехом использованные палеоэнтомологами (Жерихин, 1978, 1979; Расницын., 1988). Это связано с тем, что при расчете всех этих индексов так или иначе используются соотношения существующих, появляющихся и исчезающих таксонов. Поэтому мы приводим здесь только динамику разнообразия существующих в каждый момент семейств и родов. Но и эти данные достаточно красноречиво свидетельствуют о беспрецедентном сокращении разнообразия наземных тетрапод в ранней—средней юре.
М. Бентон (Benton, 1985—1989), независимо от нас предпринявший исследование динамики таксономического разнообразия наземных тетрапод, получил очень близкую картину.
Зоогеографическая раздробленность. Повышать стратиграфическую и таксономическую точность при глобальном исследовании динамики разнообразия тетрапод на протяжении всего времени их существования вряд ли целесообразно, по крайней мере по причинам зоогеографического порядка. Сложение таксономического разнообразия разных, географически изолированных стран с весьма контрастной фауной (рис. 1) осмысленно лишь для самых ориентировочных экологических оценок и оправдано лишь необходимостью преодолевать пространственную неполноту палеонтологической летописи, которая временами досадно велика. Например, пик разнообразия амфибий позднего карбона — ранней перми известен только для Еврамерийской зоогеографической области. В поздней перми их разнообразие резко сокращается, несмотря на добавление Гонд-вандской области, т. е. большей части суши (см. рис 1). Таким образом, сокращение разнообразия амфибий в Еврамерии было более резким, чем на глобальной картине (Каландадзе, Раутиан, 1983). В Гондване в это время уже появляются группы, потомки которых образовали раннетриасовый максимум родового разнообразия амфибий (рис. 2, б). Это свидетельствует в пользу более ранней смены разнообразия амфибий в Гондване, возможно связанной с пермо-карбоновым оледенением. Аналогичные трудности глобального анализа разнообразия встречаются во все периоды зоогеографической раздробленности (см. рис. 1).
|
|
|
Для временной окрестности интересующего нас кризиса проблема зоогеографической раздробленности сведена к минимуму. Ему предшествует зоогеографическая «пангея» позднего триаса — эпоха максимальной консолидации фаун различных континентов за весь исследованный период, уничтожившая предшествующее зоогеографи-ческое деление (Каландадзе, Раутиан, 1980, 1981, 1983, 1991).
А был ли кризис? Бедные местонахождения наземных тетрапод ранней—средней юры, к сожалению, не подвергались подробным тафономическим исследованиям. Поэтому судить о том, не является ли резкое обеднение фауны в это время следствием тафономического артефакта, как это уже предполагалось (Несов, Казнышкина, Черепанов, 1989), приходится косвенным путем. Л. А. Несов полагает, что причиной обеднения фауны мог быть гумидный климат, создающий неблагоприятные условия захоронения. Действительно, теплый и гумидный климат юры не отличался существенным контрастом в различных частях суши. Однако не настолько, чтобы отсутствовала климатическая зональность (Хэллем, 1978; Рид, Уотсон, 1981; Вахрамеев, 1988). Поэтому трудно себе представить, чтобы условия захоронения даже крупных тетрапод во всех климатических поясах были одинаково плохими. Это соображение в сочетании с широкой географической представленностью и монотонной бедностью местонахождений, расположенных в областях с заведомо отличающимся климатом (ср. Каландадзе, Раутиан, 1983 и Вахрамеев и др., 1970; Вахрамеев, 1988), свидетельствуют не в пользу предположения Л. А. Несова. Кроме того, поздняя юра с ее гораздо более богатой фауной тоже не отличалась значительной аридностью и климатическим контрастом.
|
|
|
Условия жизни в юре. Неконтрастный климат юры вряд ли мог спровоцировать глобальный экологический кризис. К таким глубоким последствиям, как в ранней—средней юре, не привели даже оледенения пермо-карбона и плейстоцена. Не было в это время и других особенно крупных физико-географических переворотов (Хэллем, 1978): особенно сильных глобальных трансгрессий или регрессий (во всяком случае, регрессия в поздней перми и трансгрессия в позднем мелу намного сильнее юрских: Борисяк, 1934; Страхов, 1948), аномально высокой вулканической активности (Рид, Уотсон, 1981) и т. п.
Синхронное сокращение разнообразия. На любом отрезке истории тетрапод, равном по длительности ранней—средней юре, можно обнаружить семейства и группы более высокого ранга, обладающие в это время максимумом разнообразия за всю историю. Эпоха рассматриваемого кризиса является единственной, когда ни одно семейство, не говоря о таксонах более высокого ранга, не дало максимума разнообразия и не ограничивало своего стратиграфического распространения только этим интервалом. Синхронное сокращение разнообразия всех групп наземных тетрапод до семейственного уровня включительно исключает объяснение этого феномена посредством конкурентного вытеснения групп в пределах самих тетрапод.
Отсутствие синхронной реакции других групп. Нелегко представить себе другую группу животных, способную столь радикально повлиять на всех наземных тетрапод. Не наблюдается в ранней— средней юре резкой смены растительности. Насекомые, четко реагирующие на смену растительности (Жерихин, 1978—1987), также не испытали в юре значительных фаунистических перемен (Расницын, 1988; Дмитриев, Жерихин, 1988). Во всяком случае, смена тех (Вахрамеев и др., 1970; Вахрамеев, 1988; Красилов, 1989) и других (Жерихин, 1978—1987) в середине мела представляется гораздо более радикальной. Испытали кризис в середине мела и наземные тетраподы (Жерихин, 1979), но существенно меньшего масштаба, чем в юре.
Космические факторы. Отсутствие ярко выраженной синхронной реакции других компонентов биоты заставляет скептически относиться к возможной космической причине юрского кризиса в сообществе наземных тетрапод, хотя такие же предположения выдвигались (Paul, 1988). Космический катаклизм вряд ли мог обладать такой высокой нсспецифичностью и одновременно избирательностью, чтобы радикально повлиять на все группы наземных тетрапод, но только на них.
Внешние факторы по отношению к фауне наземных тетрапод могли в существенных чертах определить наступление кризиса только в случае их экстраординарности и исторической нетипичности (Шмальгаузен, 1968—1983), судя по реакции наземных тетрапод. Они должны обладать глобальным и, как отмечалось, одновременно неспецифическим и высокоизбирательным действием, т. е., грубо говоря, лежать на поверхности, чего, как следует из предыдущего, не видно. Кроме того, даже если такие факторы обнаружатся, то они способны лишь спровоцировать кризис, но ответная реакция на них все равно в большей степени должна определяться свойствами реагента, в данном случае фауны, чем свойствами воздействия (Раутиан, 1988). Во всяком случае, резкая фаунистическая смена окажется возможной только тогда, когда к моменту кризиса фауна окажется «беременной» соответствующими таксономическими и экологическими новообразованиями. Все это заставляет скептически относиться к гипотезе экзогенной причины кризиса.
Симптомы экологического кризиса. Всякий истинный экологический кризис, в отличие от случаев обеднения палеонтологической летописи в силу тафономических и/или геологических причин (Ефремов, 1950; Muller, 1986) должен сопровождаться крупными преобразованиями в таксономическом составе биоты и экологической структуре сообщества. Ниже приводится расширенный (по сравнению с нашей прежней публикацией: Каландадзе, Раутиан, 1983) перечень симптомов экологического кризиса, выявленных в результате исследований биоценотического кризиса в середине мела (Родендорф, Жерихин, 1974; Жерихин, 1978—1987; Расницын, 1988) и кризиса сообщества наземных тетрапод в ранней—средней юре (Каландадзе, Раутиан, 1983, 1984).
1. Непосредственно в момент кризиса характерно резкое падение таксономического разнообразия биоты, которое связано не столько с увеличением скорости вымирания в момент кризиса, сколько с понижением скорости появления новых таксонов.
2. Появление во временной окрестности кризиса большого числа новых таксонов, в том числе высокого ранга.
3. Вымирание во временной окрестности кризиса множества прежде широко распространенных таксонов, в том числе высокого ранга.
4. Большая часть таксонов высокого (при крупном кризисе семейственного и более высокого) ранга исчезает из палеонтологической летописи или появляется в ней либо до, либо после кризиса.
5. Возрастание доли и понижение ранга таксонов, достигающих современности, во временной окрестности кризиса и в послекризисных биотах.
6. Викарные замены во временной окрестности кризиса ряда древних таксонов высокого ранга филогенетически преемственными по отношению к ним таксонами тоже высокого ранга.
7. Смена экологических доминантов во временной окрестности кризиса, выражающаяся в переходе ряда таксонов из категории редких и редчайших в категорию регулярно попадающих в палеонтологическую летопись.
8. Появление во временной окрестности кризиса большого числа короткоживущих (недолго встречающихся в палеонтологической летописи) таксонов, в том числе сравнительно высокого (при крупном кризисе главным образом семейственного) ранга.
9. Появление таксонов, регулярно попадающих в палеонтологическую летопись только во временной окрестности кризиса.
10. Выпадение в разгар кризиса из палеонтологической летописи ряда таксонов, сохраняющихся в ходе кризиса без существенных изменений.
11. Крупному экологическому кризису предшествует кризис меньшего масштаба.
12. Новая биота (сообщество), возникшая сразу после крупного экологического кризиса, оказывается недолговечной и вскоре снова ввергается в кризис, но меньшего масштаба.
13. В разгар крупного экологического кризиса широкая географическая представленность местонахождений сопровождается крайней бедностью их биоты и очень слабыми таксономическими связями.
14. Глобальному экологическому кризису предшествует истори-ко-зоогеографическая «пангея» — относительный континуум биоты всех стран, подверженных кризису.
Несомненно, ни один из этих симптомов в отдельности не гарантирует надежную диагностику экологического кризиса, хотя для «нащупывания» эпохи, перспективной для поисков кризиса, можно использовать и отдельные симптомы. На поиски, юрского кризиса, как отмечалось, нас натолкнуло сочетание 1 и 13 симптомов. Совпадение же многих, и особенно большинства симптомов, трудно убедительно объяснить при помощи иного предположения.
Временная окрестность кризиса, с которой связано большинство симптомов (2—9-й, 11—12-й, 14-й), важна по двум причинам. Во-первых, обнаружение в биоте предвестников кризиса и его длительного последствия косвенно свидетельствуют о наличии преемственности в процессах филоценогенеза и, следовательно, в пользу реальности ценотических структур, с которыми имеет дело палеонтолог (Раутиан, 1988). Историческая обусловленность течения и релаксации кризиса — важнейший показатель их эндогенной обусловленности даже в том случае, когда кризис был непосредственно спровоцирован внешней по отношению к биоте причиной. Именно поэтому сторонники большой роли внешних (например, космических) факторов стараются аргументировать положение о неожиданности кризиса для биоты («Катастрофы и история Земли», 1986; Татаринов, 1987). Во-вторых, резкое обеднение таксономического разнообразия во время кризиса (1-й симптом) резко ограничивает доступный для анализа материал. Раздвижение рамок временной окрестности кризиса позволяет благодаря преемственности частично преодолеть удручающую неполноту материала в разгар кризиса.
За окрестности ранне-среднеюрского кризиса нами условно принят весьма широкий интервал, начиная с позднего триаса и кончая поздней юрой. В таблице представлены таксоны наземных тетрапод (не ниже уровня семейства), известные из этого интервала, но их стратиграфическое распространение показано, начиная с раннего триаса и до современности.
Симптомы ранне-среднеюрского кризиса. Ниже рассматриваются проявления всех симптомов на примере юрского кризиса, кроме 11-го симптома. Последний для своего выявления потребовал более детальной проработки данных по динамике таксономического разнообразия, которая в настоящее время не завершена. Поэтому анализ этого симптома мы вынуждены отложить до следующей публикации.
1-й и 13-й симптомы уже обсуждались в начале статьи. По поводу скоростей вымирания и появления таксонов (второе положение 1-го симптома) необходимо добавить, что во временной окрестности кризиса не наблюдается существенного расхождения кривых динамики разнообразия появляющихся, исчезающих и существующих в данный момент семейств и родов. Следовательно, именно замедление скорости появления новых таксонов приводит к обеднению биоты в разгар кризиса, поскольку существовать и вымирать может только то, что уже появилось. Это дополнительно подтверждается тем, что основные волны исчезновения и появления таксонов (4-й симптом) приурочены к позднему триасу и поздней юре.
2—5 -и, 8-й и 10-й симптймы. Данные по ним легко извлекаются из табл. Резкое преобладание вымирания и появления таксонов в позднем триасе и поздней юре, т. е. до и после кризиса (4-й симптом), кроме общего обеднения фауны в ранней—средней юре, может иметь следующие причины. В процессах вымирания, Очевидно, немалую роль играет прямое или опосредованное конкурентное вытеснение, которое может иметь существенное значение только в экологически насыщенных, т. е. некризисных, сообществах. В. В. Жерихин (1978—1987) отмечал, что замедление процессов Вымирания в разгар кризиса может быть связано со снижением Конкурентного давления, обусловленным предшествующим нескомпенсированным вымиранием (1-й симптом).
Становление новых таксонов, особенно высокого ранга, связано с направленными филогенетическими процессами (Шмальгаузен, 1968—1983; Татаринов, 1976, 1987; Раутиан, 1988), которые значительно облегчает относительно стабильная биотическая среда сформированного сообщества. Однако жесткая экологическая структура последнего затрудняет нахождение условий для формирования новых адаптивных зон и связанных с ними направлений прогрессивной специализации (Жерихин, 1978—1987; Расницын, 1987, 1988). Начинающееся разрушение этой структуры накануне кризиса или ее формирование после кризиса создают относительно благоприятные, хотя и компромиссные условия таксонообразования и в первом и во втором отношении. Напротив, максимально неопределенная биотическая среда в разгар кризиса должна тормозить образование крупных таксонов. Это обстоятельство также будет способствовать выполнению условий 1-го симптома.
Характеристику 5-го симптома, кроме содержащегося в табл., дополняет обратная Лайелева кривая (выполненная в стиле В. В. Жерихина: Родендорф, Жерихин, 1974; Жерихин, 1978, 1979; Рас-ницын, 1988), которая показывает быстрое сокращение доли чисто ископаемых семейств и рост доли семейств, достигающих современности, в послекризисных, а отчасти и кризисных биотах (рис. 3). Можно считать, что доживающие доныне семейства наземных тетрапод появляются в палеонтологической летописи начиная с юры. Исключение составляет только появившееся в раннем триасе семейство Sphenodontidae, достигающее современности в виде единственного рода Sphenodon (гаттерия), распространение которого ограничено Новой Зеландией. Таким образом, это именно то исключение, которое подтверждает правило.

Появление во временной окрестности кризиса особенно большого количества короткоживущих таксонов преимущественно семейственного ранга (8-й симптом) хорошо согласуется с одним из правил Э. Д. Копа (Соре, 1904), в соответствии с которым перед моментом вымирания крупная группа несколько увеличивает свое таксономическое разнообразие, и правилом относительно высокого архаического разнообразия в рамках крупного таксона, которое, по-видимому, особенно четко формулировал О. Шиндевольф (Schindewolf, 1950). Яркое проявление обоих этих правил во временной окрестности кризиса свидетельствует об активизации таксонообразования в это время, о множестве проб и относительно небольшом числе удачных попыток сформировать долговременные направления прогрессивной специализации (Раутиан, 1988).
Короткоживушие таксоны временной окрестности кризиса, не выходящие за пределы одного отдела стратиграфической шкалы, встречаются только в позднем триасе и поздней юре. Это хорошо согласуется с положением 4-го симптома об активизации таксонообразования накануне и после кризиса и положением 1-го симптома об угнетении этого процесса в разгар кризиса.
Резкое преобладание числа таксонов, встречающихся только в позднем триасе и только в поздней юре, над таксонами, встречающимися более чем в одном отделе временнбй окрестности кризиса, но не выходящими за его пределы, свидетельствует в пользу реальности существования группы короткоживущих таксонов.
Выпадение в разгар кризиса большого числа таксонов, встречающихся до и после кризиса (10-й симптом) можно связать с резким ухудшением биотической среды в кризисных сообществах. При этом они не вымерли, но их численность и/или ареал могли сокращаться настолько, что оказывались за пределами тафономически достоверной части палеонтологической летописи.
б-й и 7-й симптомы удобно рассматривать вместе. Lepospondyli. Apoda появляются в позднем триасе, a Urodela в средней юре (если не считать сомнительного Triassurus из позднего триаса: Ивахненко, 1978). Их предполагаемые предки — Tuditanomorpha, а также другие древние группы Lepospondyli — не известны позже верхней перми (Carrol, Gaskill, 1978; Carroll, 1988). Следовательно, во временнбй окрестности кризиса Lepospondyli после длительного перерыва вновь появляются в палеонтологической летописи, причем в виде двух новых, ранее неизвестных, отрядов, обладающих ныне значительным таксономическим разнообразием и обширными ареалами: Apoda — циркумтропическим, a Urodela — почти циркумголарктическим.
Labyrinthodontia. Batrachomorpha, среди которых были предки Salientia — Dissorophidae, известны с позднего девона до раннего мела, причем с карбона до позднего триаса в качестве экологических доми-нантов. Позже были только спорадические находки: Capitosauridae в лиманном бассейне среднеюрского Тетиса (Nessov, 1988), Brachyopidae в позднеюрском озере Монголии (Шишкин, 1991) и Plagiosauridae в апт-альбе (Milner, 1989), изолированной уже в это время Австралии (рис. 1), что подчеркивает реликтовый характер находки. Древнейшие Salientia представлены изолированной находкой представители монотипического отряда Proanura в раннем триасе Мадагаскара. Первые представители современного отряда Anura, испытавшего в кайнозое фазу биологического прогресса (Северцов, 1978) и имеющего ныне огромное разнообразие и субглобальный ареал, известны начиная с ранней юры и имеют начиная с поздней юры непрерывную палеонтологическую летопись. Таким образом, во временнбй окрестности кризиса Batrachomorpha переходят в категорию палеонтологических редкостей, а в раннем мелу вымирают, Salientia, напротив, переходят в категорию экологических доминантов, а Anura появляются в разгар кризиса сразу в виде современного, хотя и реликтового семейства Leiopelmatidae. В поздней юре к нему добавляется второе, тоже современное, но шире распространенное семейство Discoglossidae, имеющее начиная с позднего мела непрерывную палеонтологическую летопись. Оба эти семейства входят в подотряд Archaeobatrachia.
Parareptilia. Черепахи (Chelonia) появляются в позднем триасе одновременно в виде архаичного подотряда Proganochelydia и самого продвинутого разнообразного и широко распространенного ныне инфраотряда Cryptodira. Первый был широко распространен в позднем триасе Европы, второй представлен единственной находкой из Северной Америки. Начиная с поздней юры оба современных инфраотряда Pleurodira и Cryptodira переходят в категорию экологических доминантов. Предполагаемые предки черепах Pareiasauria распространены и многочисленны только в поздней перми. Позже неизвестны и другие архаичные отряды подкласса Diadecfosauria, к которому М. Ф. Ивахненко (1978) относит черепах. Следовательно, перед самым кризисом после длительного перерыва этот подкласс вновь появляется в палеонтологической летописи, причем в виде нового, ранее неизвестного, отряда, современные инфраотряды которого переходят в категорию экологических доминантов сразу после кризиса. Эта последовательность событий сходна с таковой для Lepospondyli.
Squamata. Древний подотряд ящериц — Eolacertilia экологически доминирует в поздней перми—позднем триасе, хотя последние его представители известны из позднего мела. Происходящий от него подотряд Lacertilia с поздней юры имеет непрерывную палеонтологическую летопись, а ныне — огромное разнообразие и субглобальный ареал. Он сразу появляется в виде современных инфраотрядов:
Gekkota, Sdncomorpha (с современным семейством Sdncidae) и Anguimorpha (с современными надсемействами Anguioidea и Varanoidea). Таким образом, древнейший подотряд ящериц в результате кризиса переходит в категорию палеонтологических редкостей, а новый, ранее неизвестный, подотряд после кризиса переходит в категорию экологических доминантов. Varanoidea появляются в палеонтологической летописи в виде примитивного семейства морских обитателей Aigialosauridae. Следовательно, учитывая несомненно наземное становление надсемейства, его представители сначала становятся экологическими доминантами в море (которого не коснулся интересующий нас кризис) и лишь в позднем мелу занимают это положение в сообществах суши.
Archosauria. Thecodontia — экологические доминанты триаса — вымирают к концу этого периода. Им на смену в позднем триасе приходят происходящие от них экологические доминанты позднего мезозоя: Crocodylia, Pterosauria и Dinosauria.
Крокодилы. Древнейший подотряд Profosuchia вымирает лишь в позднем мелу, но уже в ранней юре порождает более продвинутый и разнообразный подотряд Mesosuchia. Первые известные представители последнего — морские обитатели, хотя формирование подотряда, несомненно, происходило на континенте. Наземные Mesosuchia неизвестны до поздней юры. Следовательно, появившись в разгар кризиса, они сначала становятся экологическими доминантами в море и только после кризиса занимают это положение в сообществах суши, которое удерживают до конца мезозоя. Их разнообразие резко сокращается в течение палеогена в связи с формированием крупных специализированных хищных млекопитающих. Последние несомненные представители тоже сухопутные (Sebecidae) доживают до миоцена в изолированной (см. рис. 1) Южной Америке (Раутиан, Каландадзе, 1989), что подчеркивает реликтовый характер группы в это время.
Птерозавры. Древний подотряд Rhamphorhynchoidea экологически доминирует в позднем триасе—ранней юре и вымирает в поздней юре, где его в роли экологических доминантов сменяет подотряд Pterodactyloidea, возникший в средней юре. Таким образом, все важнейшие события в филогенетической и экологической истории птерозавров (кроме их вымирания в позднем мелу) связаны с эпохой кризиса и его временной окрестностью.
Динозавры в позднем триасе уже представлены всеми группами подотрядного ранга, кроме Maniraptora, которые известны начиная с поздней юры. Достоверных остатков Fabrosauria — древнейших Thyreophora — не известно позже раннего мела. В средней юре появляются происходящие от них Stegosauria и Ankylosauria, причем первые экологически доминируют в поздней юре—раннем мелу, а вторые — в позднем мелу. Pachycephalosauria начиная с поздней юры сменяют своих предков Heterodontosauria, экологически доминировавших в позднем триасе, но последняя находка которых относится к средней юре Китая. Проблематичные остатки примитивных Hypsilophodontia известны из позднего триаса и средней юры. Начиная с поздней юры в категорию экологических доминантов переходят Iguanodontia. Среди Saurischia экологическими доминантами позднего триаса были Prosauropoda и Podokesauridae, стратиграфическое распространение которых ограничивается этим периодом. Начиная с ранней юры им на смену приходят происходящие от них соответственно Cetoisauria и Carnosauria, экологически доминировавшие до конца мезозоя. Начиная со средней юры к ним добавляются Coelurosauria, которые происходят от Podokesauridae. Maniraptora (они происходят от примитивных динозавров, по уровню специализации соответствующих примерно триасовым Prosauropoda и Podokesauridae, и порождают уже в поздней юре примитивных птиц Praeornithes (Раутиан, 1978) и Archaeoornithes), очевидно, в поздней юре лишь начинают свою экологическую экспансию, развернувшуюся в мелу, а возникнуть должны были в конце триаса — начале юры. Таким образом, большая часть крупнейших филогенетических и экологических вобытий в истории динозавров тоже связана с эпохой кризиса и ее временной окрестностью.
Theromorpha. Эпоха экологического доминирования Synapsida завершается в позднем триасе, хотя последняя находка Tritylodontidae относится к средней юре Китая. Их филогенетические потомки — млекопитающие — появляются в позднем триасе, причем сразу представлены оба современных подкласса: Prototheria и Theria. Morganucodonta широко представлены только в позднем триасе, хотя последние их находки относятся к раннему мелу. В средней юре от них происходят Eutriconodonta, экологически доминировавшие в поздней юре—раннем мелу, но дожившие до позднего мела. Средней—поздней юрой ограничено распространение Docodonta. В поздней юре появляются Multituberculata, экологически доминировавшие до эоцена включительно и испытавшие пик родового разнообразия в палеоцене (Раутиан, 1980). Ни одна группа Pantotheria, кроме позднеюрских Dryolestia, не имеет ярко выраженной эпохи экологического доминирования. Тем не менее и здесь можно наметить определенную смену. Kuehneotheriidae имеют напрерывную палеонтологическую летопись с позднего триаса до поздней юры, а происходящие от них Spalacotheria известны с ранней юры, Eupantolheria — со средней, Shuotheridia ограничены поздней юрой. Таким образом, все существенные филогенетические и экологические события в истории древних групп млекопитающих — Pantotheria и Prototheria (кроме Multituberculata, имеющих длинную историю) — связаны с эпохой кризиса и его временной окрестности.
Какой кризис крупнее? В целом можно сказать, что после ранне-среднеюрского кризиса, т. е. начиная с поздней юры, структура биоты наземных тетрапод мелкого размерного класса становится принципиально кайнозойской: хвостатые и безхвостые амфибии, бокошейные и скрытошейные черепахи, ящерицы современных инфраотрядов и даже семейств, птицы и млекопитающие (последние две группы, правда, представлены архаичными формами). Все эти группы, возможно, кроме птиц, уже в поздней юре вышли в категорию экологических доминантов. Самой массовой группой мезозойских млекопитающих являются Multituberculata, обладающие адаптацией принципиально грызунового типа, а грызуны — самый массовый отряд современных млекопитающих.
В крупном размерном классе — иная картина. В течение всего позднего мезозоя здесь господствовали архозавры: динозавры, птерозавры и крокодилы. Именно переменой главных действующих лиц в крупном размерном классе и обусловлен кризис на рубеже мела и палеогена. Вымирание большинства крупных архозавров открыло членам уже принципиально кайнозойского сообщества мелких тетрапод возможность освоения адаптивных зон крупных животных. Этой возможностью с большим или меньшим успехом воспользовались все перечисленные выше группы первично мелких тетрапод, кроме, пожалуй, бесхвостых амфибий.
Все это свидетельствует о более масштабной фаунистической смене в ранней—средней юре по сравнению с таковой на рубеже мела и палеогена. При этом результаты первой из них в значительной степени определили характер второй. Последнее заставляет со скепсисом относиться (Татаринов, 1987) к представлениям об определяющей роли внешних (например, космических) причин и в процессах мел-палеогеновой смены, по крайней мере у наземных тетрапод.
Фаунистической смены, подобной по масштабам юрской, мц не знаем и во всем интервале с карбона до триаса. По-видимому, крупнейшим известным событием этого интервала был экологический кризис на рубеже перми и триаса. Ему предшествовал момент насыщения биоценозов суши животными с рептильным уровнем организации (см. первый максимум разнообразия рептилий в поздней перми на рис. 2), в котором решающую роль играли зверообразные (Каландадзе, Раутиан, 1983). Главными событиями, обусловившими кризис на рубеже перми и триаса, были: 1) внедрение быстро передвигающегося хищника (Thecodontia) в принципиально тихоходную фауну тетрапод крупного размерного класса и 2) интенсивное освоение рептилиями водных местообитаний. Первое из них будет освещено в подготавливаемой А. Г. Сенниковым диссертации, а второе было коротко рассмотрено нами (Каландадзе, Раутиан, 1983).
Таким образом, ранне-среднеюрский экологический кризис сопровождался крупнейшей фаунистической сменой за всю палеонтологически документированную историю наземных тетрапод.
9-й симптом ярче всего выражают Leiopelmatidae — современное семейство, известное в ископаемом состоянии только в ранней и поздней юре. Scelidosauridae известны в позднем триасе—ранней юре, а за пределами временней окрестности кризиса известна всего одна находка (Minmi) из апт-альба изолированной в это время Австралии (см. рис. 1), что дополнительно подчеркивает реликтовый характер группы в это время. Morganucodonta имеют сплошную палеонтологическую летопись в позднем триасе—средней юре, а за пределами временной окрестности кризиса представлены одной ран-немеловой находкой. Ранние Multituberculata (Plagiaulacida) представлены только в поздней юре и базальном раннем мелу. Большая часть находок Kuehneotheriidae относится к позднему триасу—ранней юре. До поздней юры они сохраняют непрерывную палеонтологическую летопись, а за пределами временной окрестности кризиса имеется только одна находка из позднего мела Северной Америки. Большая часть находок Peramuridae относится к средней—поздней юре, a Spalacotheriidae — только к поздней юре. За пределами временной окрестности кризиса оба эти семейства известны лишь по спорадическим находкам в раннем—позднем мелу.
12-й симптом. Выраженный пик таксономического разнообразия в поздней юре и последующий его спад в раннем мелу сопровождался заметной фаунистической сменой (рис. 2, б), очевидно, отражающей изменения в экологической структуре сообщества наземных тетрапод (Жерихин, 1979). Однако масштаб этого события значительно скромнее ранне-среднеюрского. Предварительные данные более детального анализа (который не входит в задачи этой статьи) показывает, что выраженный рост разнообразия, особенно родового, относится лишь к верхней части поздней юры (киммеридж—портланд). Оксфордская фауна, по-видимому, еще носила кризисный характер. Последующее снижение разнообразия относится к неокому, а в апт-альбе уже начинается новый рост разнообразия, переходящий в поздний мел. Последствия этого небольшого кризиса имеют в отличие от раннесреднеюрского выраженные региональные особенности. Последнее связано с зоогеогра-фической раздробленностью, характерной для второй половины мела (рис. 1). Таким образом, уже сейчас можно довольно уверенно говорить об экологической природе события в неокоме и тем самым о недолговечности первого послекризисного сообщества наземных тетрапод.
14-й симптом. Глобальный экологический кризис предполагает необходимость глобальных экологических взаимодействий, когда принципиально единой биоте соответствует принципиально единое глобальное сообщество. Важнейшей причиной, нарушающей единство биоты наземных тетрапод, является морская изоляция (Элтон, I960; Каландадзе, Раутиан, 1983; Wallace, 1876). И действительно, глобальному экологическому кризису в ранней—средней юре предшествует эпоха зоогеографической «пангеи» позднего триаса, которая, по-видимому, продолжалась и в начале юры (см. рис. 1). В это время наблюдалась максимальная консолидация всех фаун наземных тетрапод за весь период их палеонтологически документированной истории (Каландадзе, Раутиан, 1980, 1981, 1983, 1991). Активный обмен между фаунами, нивелировка прежнего зоогеографического деления и заселение всех палеонтологически охарактеризованных стран однородной фауной наземных тетрапод, сложившейся в Гон-дванской зоогеографической области, способствовали общему обеднению фауны позднего триаса за счет вымирания большинства специфических для Еврамерийской области таксонов. Единственным известным исключением являются Lepospondyli — еврамерийский по своему происхождению элемент, доживший до нашего времени в виде Apoda и Urodela. Все перечисленные обстоятельства способствовали глобальному проявлению ранне-среднеюрского кризиса. Однако ни одно из них в отдельности, ни все они вместе как таковые не могли спровоцировать кризис и тем более не могут дать" удовлетворительного объяснения синхронности перемен, охвативших все крупные группы наземных тетрапод в период кризиса. Таким образом, временное соседство зоогеографической «пангеи» позднего триаса и экологического кризиса в ранней—средней юре можно считать необходимым, но недостаточным условием последнего.
Проведенный анализ юрского кризиса вряд ли оставляет сомнения в реальности этого события, его исторической подготовленности и длительном последействии. Однако это всего лишь необходимые условия постановки задачи по систематическому исследованию юрского кризиса.
Причина юрского кризиса, сопровождавшегося снижением таксономического разнообразия всех групп наземных тетрапод до семейственного уровня включительно (с. 64), должна прежде всего объяснить эту синхронизацию. Самым простым является предположение о синхронизирующем влиянии глобального ухудшения условий жизни наземных тетрапод, вызванного внешней по отношению к ним причиной. Выше (с. 64—65) было показано, с какими трудностями встречается это предположение и почему оно не представляется правдоподобным.
Наиболее экзотическим является предположение о чисто внутренней (автогенетической) причине. Однако рассмотрим и его. Причину кризиса можно искать в филогенетическом единстве тетрапод. Такое предположение хорошо объясняет синхронность реакции тетрапод в период кризиса и отсутствие такой реакции других элементов биоты. Но в этом случае весьма древняя дивергенция главных стволов тетрапод, представленных во временной окрестности кризиса, заставляет предположить сохранение унаследованного ритма прохождения основных фаз биологического прогресса и регресса. Последнее заведомо ложно, поскольку удивительная синхронность реакции наземных тетрапод в ранней—средней юре в этом случае должна была захватить и морских тетрапод, а также должна была наблюдаться в порядке правила на всем интервале, по крайней мере с карбона до юры включительно. Не наблюдается ни то, ни другое. Кроме того, гиперболизация роли внутренней исторической причины неизбежно ведет к признанию отсутствия заметн,ой связи динамики таксономического разнообразия с экологией. А это противоречит многим правдоподобным реконструкциям филоценогенеза, в том числе выполненным в Палеонтологическом институте АН СССР (Жерихин, 1978—1987; Каландадзе, Раутиан, 1979, 1983; Раутиан, 1980; Расницьга, 1988; Раутиан, Каландадзе, 1989).
Наиболее реалистическим подходом к решению юрской и аналогичных проблем нам представляется гипотеза наличия специфической экологической организации наземных тетрапод, образующих не только фауну, как совокупность таксонов, населяющих некоторое пространство, но и сообщество — коадаптивный комплекс (Длусский, 1981) в пределах биоценоза, который, в свою очередь, является продуктом длительной сопряженной эволюции. Природа кризисных ситуаций в соответствии с этой гипотезой заключается в нарастании противоречий в стратегиях эволюции коадаптивного комплекса, с одной стороны, и входящих в него филогенезов — с другой. Отсутствие реакции всего биоценоза в этом случае свидетельствует в пользу значительной автономности сообщества наземных тетрапод от остальной биоты, а синхронность его реакции — в пользу его значительной функциональной целостности, потеря которой сообществом в целом и выразилась в кризисной ситуации. В пользу существования автономного экологического комплекса наземных тетрапод свидетельствует, в частности, тот факт, что главные события их экологической эволюции, кратко рассмотренные в наших прежних публикациях (Каландадзе, Раутиан, 1979, 1983; Раутиан, Каландадзе, 1989), удовлетворительно объясняются без апелляции к другим компонентам биоты или физическим факторам среды.
Ниже предлагается модель филоценогенеза, в которой, в частности, закономерно, хотя и не часто, возникают ситуации крупных экологических кризисов.
|
|
|
|
|
Дата добавления: 2015-05-29; Просмотров: 482; Нарушение авторских прав?; Мы поможем в написании вашей работы!