КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Для того, чтобы направлять белки в тилакоидную мембрану хлоропластов, необходимы два сигнальных
8-18
пептида [27]
Транспорт белков в хлоропласти во многих отношениях напоминает транспорт в митохондрии: и тот, и другой процесс происходят после трансляции, и тот, и другой нуждаются в энергии, и в том, и в другом случае используются гидрофильные N-концевые сигнальные пептиды, которые затем удаляются. Однако имеется по крайней мере одно важное отличие: в митохондриях для проведения этого транспорта используется еще электрохимический градиент на их внутренней мембране. В хлоропластах же, где электрохимический градиент имеется не на внутренней мембране, а на мембране тилакоида (см. гл. 7), по-видимому, для транспорта через внешнюю двумембранную оболочку в качестве источника энергии используется только гидролиз АТР.
Сигнальные пептиды для переноса белков в хлоропласты напоминают раннее описанные пептиды для импорта в митохондрии. Но в растительных клетках имеются и митохондрии, и хлоропласты, и соответственно, белки должны «выбирать» между ними. Например, в растительных клетках бактериальный фермент, присоединенный (методами генной инженерии) к N-концевой последовательности митохондри-
ального белка, направляется в митохондрии. Тот же белок, связанный
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
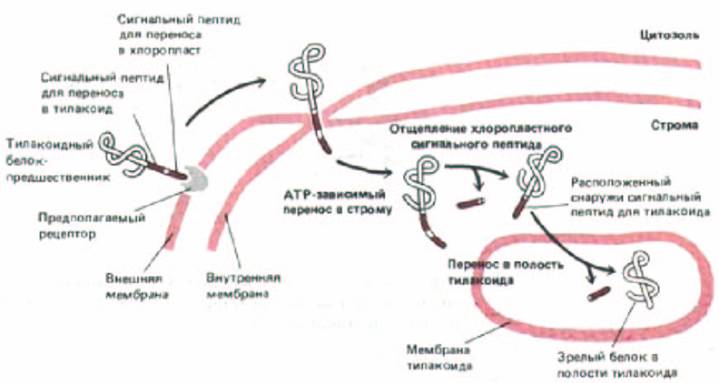
Рис. 8-31. Для переноса белков в полость (тилакоидное пространство) тилакоида в хлоропластах требуется два сигнальных пептида, такой перенос осуществляется в две стадии. Полипептнд-предшественник содержит N-кондевой сигнальный пептид для переноса в хлоропласт, за которым непосредственно следует тилакоидный сигнальный пептид. Хлоропластный сигнальный пептид вызывает перенос белка в строму через точки контакта в мембране (см. рис. 7-73); механизм этого переноса сходен с механизмом переноса в митохондриаль-ный матрикс (рис. 8-29). Затем сигнальный пептид отрезается, открывая тилакоидный сигнальный пептид, который вызывает перенос через мембрану тилакоида. Этот процесс имеет много общего с перемещением белков в ЭР.
с N-концевой последовательностью белка хлоропластов, оказывается в хлоропластах.
Хлоропласты содержат еще один окруженный мембраной компартмент - тилакоид. Множество белков хлоропластов, включая белковые субъединицы фотосинтетической системы и АТР-синтетазы, импортируются в мембрану тилакоида из цитозоля. Подобно некоторым митохондриальным белкам-предшественникам, эти белки доставляются к месту назначения в два этапа. Сначала они проникают через двумембранную оболочку в матрикс хлоропласта (называемый стромой), а затем переносятся в мембрану тилакоида (или сквозь нее в тилакоидное пространство). Предшественники этих белков имеют кроме N-концевого сигнального пептида для хлоропластов еще гидрофобный тилакоидный сигнальный пептид. После того, как белок с помощью N-концевого сигнального пептида проникает в строму, этот пептид удаляется стро-мальной протеазой (аналогичной протеазе матрикса митохондрий). В результате открывается тилакоидный сигнальный пептид, который затем инициирует транспорт через мембрану тилакоида (рис. 8-31). Как и в митохондриях, этот второй этап служит для встраивания собственных белков хлоропласта в мембрану тилакоида; необходимый для этого белок-транслокатор, вероятно, происходит от бактериального предка хлоропластов.
Заключение
Большинство белков проникает в митохондрии и хлоропласты из цитозоля сходным образом. Этот механизм был наиболее хорошо изучен для митохондрий, особенно у дрожжей. Белок переносится в матрикс митохондрии через зоны слипания внешней и внутренней мембран. Для этого переноса требуется гидролиз АТР, а также электрохимический градиент на внутренней мембране. Транспортируемый белок разворачивается, когда пересекает мито хондриальные мембраны. В митохондрии или хлоропласты переносятся только те белки, которые содержат специфический сигнальный пептид. Этот сигнальный пептид обычно расположен на N-конце молекулы белка и отрезается после переноса ее внутрь органеллы. На втором этапе транспорта белок может переноситься во внутреннюю мембрану. Для этого он должен иметь еще гидрофобный сигнальный пептид; этот пептид открывается после удаления первого сигнала. В случае хлоропластов для переноса белков из стромы в тилакоид также требуется второй сигнальный пептид.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
8.5. Пероксисомы [28]
Перокснсомы (называемые также микротельцами) во многом отличаются от митохондрий и хлоропластов. Прежде всего, они окружены только одной мембраной и не содержат ДНК и рибосом. Поскольку пероксисомы не имеют собственного генома, все их белки должны поставляться из цитозоля. В этом отношении пероксисомы напоминают ЭР: они являются самовоспроизводящейся мембранной органеллой, существующей без своего собственного генома.
Прежде чем рассматривать биосинтез пероксисом, мы остановимся на функциях этого пестрого семейства органелл. Хотя пероксисомы есть во всех эукариотических клетках, их функции сильно различаются в клетках разных типов.
В начале 60-х гг. было показано, что главным источником по крайней мере трех окислительных ферментов - оксидазы D-аминокислот, уратоксидазы и каталазы - являются самостоятельные органеллы диаметром около 0,5 мкм. У млекопитающих пероксисомы этого размера встречаются в основном в печени. На электронных микрофотографиях их можно различить по «кристалловидной» сердцевине, состоящей из уратоксидазы (рис. 8-32). Позже, когда была разработана гистохимическая окраска на каталазу - фермент, составляющий до 40% общего белка пероксисом, было показано, что пероксисомы имеются во всех клетках. В большинстве клеток пероксисомы мельче (0,15-0,25 мкм в диаметре), чем в клетках печени.
Подобно митохондрии, пероксисома - это один из главных центров утилизации кислорода в клетке. Существует гипотеза, согласно которой пероксисома представляет собой остаток древней органеллы, выполняющей у примитивных предков эукариотических клеток все функции метаболизма кислорода. Когда в атмосфере начал накапливаться кислород, производимый фотосинтезирующими бактериями, вероятно, он был токсичен для большинства клеток. Пероксисомы могли служить для снижения концентрации кислорода в клетках, одновременно используя его химическую активность для проведения важных окислительных реакций. В соответствии с этой точкой зрения последующее появление митохондрий сделало пероксисомы в значительной мере ненужными, так как многие реакции, ранее протекавшие в пероксисомах без производства энергии, теперь с помощью окислительного фосфорилирования были сопряжены с образованием АТР. Таким образом, окислительные реакции, протекающие в современных клетках - это, возможно, те реакции, которые остались необходимыми, несмотря на появление митохондрий.

Рис. 8-32. Электронная микрофотография трех пероксисом в клетке печени крысы. Паракристалличес-кие электроноплотные включения-это фермент уратоксидаза. (С любезного разрешения Daniel S. Friend.)
8.5.1. Пероксисомы используют в реакциях окисления молекулярный кислород и перекись водорода [29]
Пероксисомы получили такое название благодаря тому, что обычно в их состав входит один или более ферментов, использующих молекулярный кислород для отщепления атомов водорода от некоторых органических субстратов (обозначенных здесь R) в окислительной реакции с образованием перекиси водорода (Н2О2):
RH2 + О2 → R + Н2О2
Каталаза использует Н2О2, образованную другими ферментами в пероксисоме, для окисления множества субстратов - например, фенолов, муравьиной кислоты, формальдегида и спирта -с помощью «окислительной» реакции Н2О2 + R'H2 → R' + 2Н2О. Этот тип окислительных реакций особенно важен в клетках печени и почек, пероксисомы
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
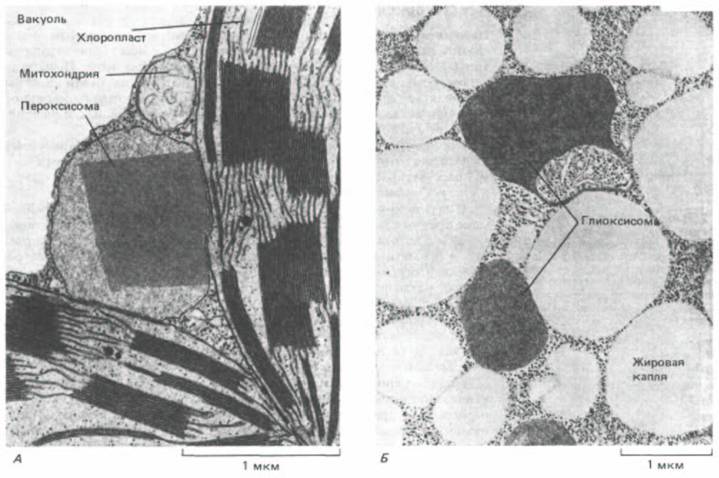
Рис. 8-33. Электронная микрофотография двух типов пероксисом, встречающихся в растительных клетках. А. Пероксисома с паракристаллической сердцевиной в клетке мезофилла листа табака. Полагают, что ее тесная связь с хлоропластами облегчает обмен материалами между этими органеллами, происходящий при фотодыхании. Б. Пероксисомы в жирозапасающей клетке семядоли семени томата, 4 дня после прорастания. Здесь пероксисомы (глиоксисомы) ассоциированы с липидными тельцами, в которых запасается жир, что отражает их центральную роль в мобилизации жиров и глюконеогенезе при прорастании семян. (А-с любезного разрешения P. Gruber и Е. Newcomb; Б с любезного разрешения S. Frederick и Е.
Newcomb.)
которых обезвреживают множество ядовитых веществ, попадающих в кровоток. Почти половина этанола, который мы выпиваем, окисляется до ацетальдегида этим способом. Кроме того, когда в клетке накапливается излишек Н2О2, каталаза превращает ее в Н2О(2Н2О2 → 2Н2О + О2).
Пероксисомы представляют собой необычно разнообразные орга-неллы, содержащие в различных клетках даже одного и того же организма сильно различающиеся наборы ферментов. В некоторых случаях в зависимости от условий изменяются размеры пероксисом. Например, клетки дрожжей, растущие в среде, содержащей сахар, имеют маленькие пероксисомы. Однако, если эти клетки выращивать на среде с метанолом, в них появляются большие пероксисомы, окисляющие метанол; если в среде есть жирные кислоты, в клетках появляются большие пероксисомы, в которых жирные кислоты расщепляются до ацетил-СоА.
Особенно важную роль пероксисомы играют в растительных клетках. У растений хорошо изучены два очень различных типа пероксисом.
Одни из них обнаруживаются в листьях (рис. 8-33, А). Эти пероксисомы катализируют окисление побочного продукта реакции, в которой СО2
превращается в углевод (такой окислительный процесс называют фотодыханием, т. к. в нем используется О2 и освобождается СО2). Другой тип пероксисом встречается в прорастающих семенах (рис. 8-33, Б). Они служат здесь для превращения жирных кислот, запасенных в липидах семян, в сахара, необходимые для роста молодого растения. Поскольку это превращение жиров в сахара происходит в серии реакций, известных под названием глиоксилатного цикла, такие пероксисомы называют еще
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
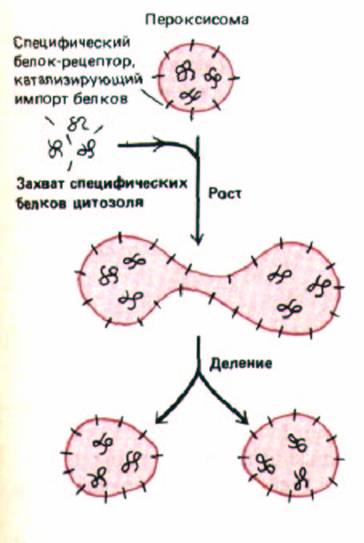
Рис. 8-34. Модель сборки пероксисом. Мембрана пероксисом содержит специфические белки-рецепторы для импорта. Все белки пероксисомы, включая новые копии этих рецепторов, синтезируются рибосомами в цитозоле и затем переносятся из цитозоля в пероксисому. Следовательно, пероксисомы образуются только из предсуществовавших пероксисом в процессе их роста и деления; подобно митохондриям и хлоропластам, они непрерывно импортируют новые компоненты из цитозоля.
глиоксисомами. В глиоксилатном цикле две молекулы ацетил-СоА, образованные в результате расщепления жирной кислоты в пероксисоме, используются для образования янтарной кислоты, которая покидает пероксисому и превращается в глюкозу. В клетках животных глиокси-латный цикл отсутствует, и поэтому они не способны превращать жирные кислоты, содержащиеся в жирах, в углеводы.
|
|
Дата добавления: 2015-08-31; Просмотров: 1857; Нарушение авторских прав?; Мы поможем в написании вашей работы!