КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Кооперативно связывающиеся кластеры белков-регуляторов могут передаваться непосредственно от
|
|
|
|
10-19
родителей к потомкам [26]
Существует несколько объяснений устойчиво наследуемой картины экспрессии генов. Одно из них основано на том, что с определенным участком хроматина кооперативно связывается множество копий регуляторного белка. Если этот белковый кластер остается соединенным с ДНК во время репликации, то часть его получает «в наследство» каждая из дочерних молекул ДНК. Поскольку связывание данного белка с ДНК носит кооперативный характер, унаследованная часть белкового кластера будет инициировать присоединение дополнительных белковых субъединиц, что в результате обеспечит реконструкцию всего кластера. Таким образом, данный функциональный статус гена наследуется прямо и непосредственно с помощью прочно связанных с ДНК хромосомных белков (рис. 10-35). В принципе подобные непосредственно наследуемые белковые кластеры могут поддерживать индивидуальные гены в постоянно включенном или постоянно выключенном состоянии.
В настоящее время еще нет убедительных доказательств в пользу описанного здесь механизма генетической регуляции, однако имеются некоторые примеры, свидетельствующие о том, что этот механизм может иметь большое значение.
10.3.8. В клетках высших эукариот гетерохроматин содержит особым образом конденсированные области ДНК [27]
Регуляторные белки, связывающиеся с определенными последовательностями ДНК в эукариотических клетках, должны взаимодействовать не просто с молекулой ДНК, как у бактерий, а с ДНК, которая на всем своем протяжении связана с нуклеосомами.
Необходимость транскрибировать ДНК в составе хроматина несомненно усложняет контроль транскрипции, однако о том, как действуют соответствующие механизмы, известно очень мало. С уверенностью можно утверждать лишь то, что у эукариот изменения в упаковке ДНК влияют на экспрессию генов. Как отмечалось выше, сайленсер, регулирующий транскрипцию у дрожжей, каким-то образом «закрывает» участки хроматина, расположенные с ним по соседству, и делает их недоступными для транскрипции и для воздействия эндонуклеазы (см. разд. 10.3.4).
|
|
|
Однако задолго до открытия этого явления изучение клеток высших эукариот продемонстрировало существование гораздо более сильно закрытого хроматина; при этом в его структуре наблюдались видимые изменения.
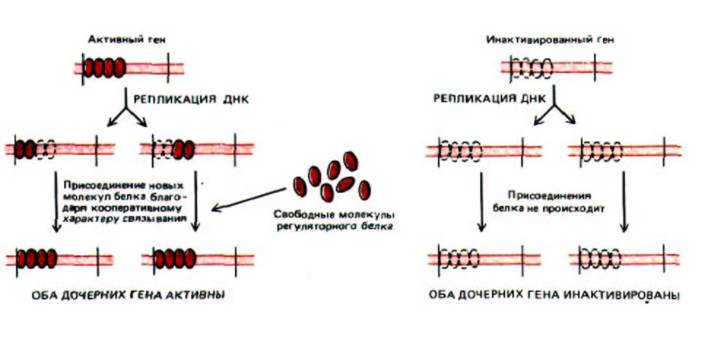
Рис. 10-35. Механизм, объясняющий прямое наследование состояния экспрессии гена в ходе репликации ДНК. Согласно этой гипотетической модели, части кооперативно связанного хластера белков-регуляторов непосредственно переносятся от родительской спирали ДНК к обеим дочерним молекулам. Унаследованный белковый кластер способствует тому, что каждая из дочерних спиралей ДНК связывает дополнительные копии тех же регуляторных белков. Поскольку связывание является кооперативным (см. рис. 9-15), ДНК, синтезировавшаяся на такой же родительской спирали, но без присоединенных регуляторных белков, не будет их связывать. Если присоединенный белок-регулятор выключает транскрипцию гена, то инактивированное состояние гена будет непосредственно наследоваться, как это происходит в случае инактивации Х-хромосомы. Если связанный белок-регулятор включает транскрипцию гена, будет наследоваться активное состояние гена.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
Изучение хромосом под световым микроскопом, проводившееся в 30-е годы, показало, что некоторые их участки не способны деконденсироваться в интерфазе и, по-видимому, сохраняют свое высоко конденсированное состояние, характерное для метафазных хромосом (см.
|
|
|
I разд. 9.2.2). Эти области были названы гетерохроматином в отличие от остального хроматина в интерфазном ядре, который носит название эухроматина. Некоторые участки хромосом конденсируются в гетерохроматин во всех клетках организма. В митотических хромосоми человека такой конститутивный гетерохроматин локализуется вокруг центромер и при специальном окрашивании легко выявляется в виде темноокрашенных зон (рис. 10-36). У некоторых других млекопитающих конститутивный гетерохроматин обнаруживается и в определенных зонах в плечах хромосом. В интерфазе участки конститутивного гетерохроматина могут агрегировать, образуя хромоцентры (см. рис. 9,43). У млекопитающих число и расположение таких хромоцентров зависит от типа клетки и стадии развития. Большинство областей конститутивного гетерохроматина содержит относительно простые тандемно-повторяющиеся последовательности, называемые сателлитной ДНК. Эти многократно повторяющиеся участки не транскрибируются, и их функция, равно как и функция формируемых ими в интерфазе конденсированных структур, остается неясной.
Некоторые области ДНК в интерфазе конденсируются и формируют гетерохроматин лишь в определенных клетках. Полагают, что эти области также не транскрибируются, однако они не состоят из простых последовательностей. Общее количество такого факультативного гетерохроматина заметно варьирует в различных клетках: в эмбриональных клетках его совсем немного, тогда как высокоспециализированные клетки отличаются высоким его содержанием. Можно предположить, что по мере развития клетки все больше и больше генов становятся неактивными вследствие того, что их ДНК приобретает конденсированную конформацию, которая не позволяет генам взаимодействовать с белками-активаторами. Большая часть данных о факультативном гетерохроматине получена при изучении инактивации одной из двух Х-хромосом в клетках самок млекопитающих.
10.3.9. Инактивация Х-хромосомы наследственно предопределена [28]
Все клетки женских особей млекопитающих имеют две Х-хромосомы, а клетки мужских организмов - одну Х- и одну Y-хромосому.
|
|
|
Предполагают, что двойная доза продуктов генов, входящих в состав Х-хромосомы, летальна для организма; возможно, именно поэтому в женских клетках возник специальный механизм, ответственный за то, что одна из двух Х-хромосом постоянно находится в инактивированном состоянии. У мышей такая инактивация происходит между третьим и шестым днем эмбрионального развития: в каждой клетке женской особи с равной вероятностью одна или другая Х-хромосома конденсируется и образует гетерохроматин. Такие конденсированные хромосомы можно увидеть в световой микроскоп: в интерфазе они представляют собой четко оформленные структурные образования, называемые тельцами Барра, которые локализованы в окрестностях ядерной мембраны. Они реплицируются в поздней S-фазе; большая часть составляющей их ДНК не транскрибируется ни в одной из дочерних клеток. Поскольку инактивированная Х-хромосома устойчиво наследуется, каждый женский организм имеет мозаичное строение в том смысле, что он образован клональными группами клеток: примерно в половине групп активна Х-хромосома, унаследованная по материнской линии (Хм), а в другой

Рис. 10-36. Хромосомы человека в метафазе. Специальная техника окрашивания позволяет выявить области конститутивного гетерохроматина (темные участки). Некоторые хромосомы помечены цифрами и буквой. (С любезного разрешения James German.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
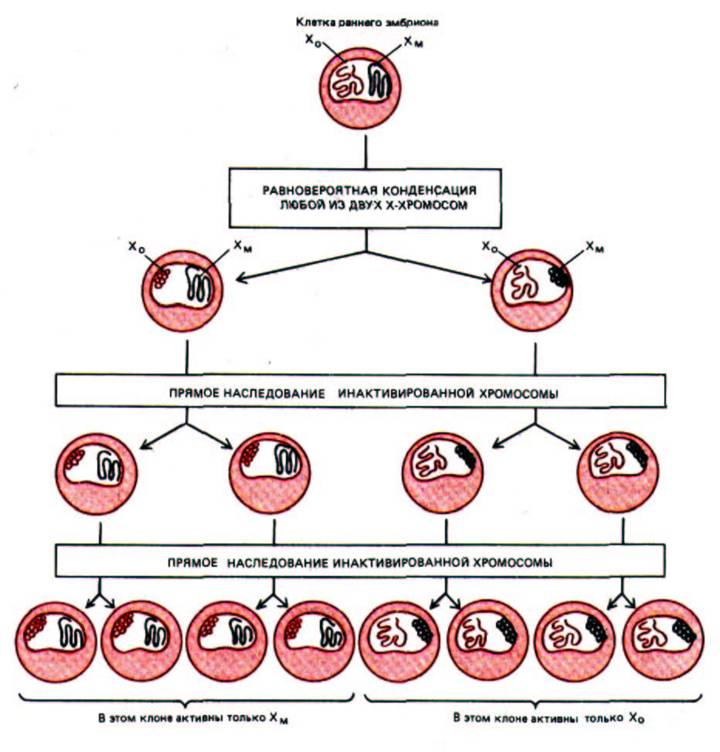
Рис. 10-37. Схема, иллюстрирующая клональное наследование конденсированной неактивной Х-хромосомы, которое имеет место у самок млекопитающих.
половине - Х-хромосома, унаследованная от отца (Х0). Иными словами, клетки, в которых экспрессируется Хм или, наоборот, Х0, расположены во взрослом организме небольшими кластерами, что отражает стремление сестринских клеток оставаться в тесном контакте друг с другом в процессе эмбрионального развития и роста организма, в то время как на ранних стадиях развития происходит некое перемешивание (рис. 10-37).
|
|
|
Процесс конденсации, в ходе которого образуется гетерохроматин X-хромосомы, имеет тенденцию распространятся вдоль хромосомы.
Это было продемонстрировано в экспериментах с мутантными особями, в клетках которых одну из Х-хромосом присоединили к концу аутосомы (неполовой, соматической хромосомы). В таких мутантных клетках участки аутосом, граничащие с инактивированной Х-хромосомой, часто конденсировались в гетерохроматин, что сопровождалось наследуемой инактивацией содержащихся в них генов. Полученные данные позволяют предполагать, что инактивация Х-хромосом - это кооперативный процесс, который можно рассматривать как «кристаллизацию», распространяющуюся из центра кристаллизации, расположенного на Х-хромосоме. После завершения конденсации хроматина такая конденсация наследуется в ходе всех последующих репликаций ДНК благодаря механизму, аналогичному тому, который представлен на рис. 10-35.
Конденсированная хромосома может вновь стать активной при формировании половых клеток. Таким образом, в ДНК, входящей в состав этой хромосомы, не происходит никаких изменений.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
10.3.10. Гены дрозофилы могут выключаться благодаря наследуемым свойствам структуры хроматина [29]
Феномен, аналогичный инактивации Х-хромосомы у самок млекопитающих, имеет место и у дрозофилы. Для его изучения весьма эффективными оказались генетические методы. У мух, несущих хромосомные перестройки, в результате которых середина гетерохроматиновой области переносится в эухроматин, эухроматиновые гены, оказавшиеся поблизости от гетерохроматина, инактивируются. Ситуация аналогична присоединению аутомосомы к неактивной Х-хромосоме у млекопитающих; инактивация в обоих случаях происходит одинаково: зона инактивации распространяется от точки разрыва хромосомы и захватывает один или несколько генов. Скорость «эффекта распространения» в разных клетках различна, но зона инактивации, возникшая в эмбриональной клетке, стабильно наследуется всеми последующими поколениями клеток (рис. 10-38).
Изучение описанного эффекта у дрозофилы показало, что скорость распространения зоны инактивации меньше у мух, геном которых содержит дополнительный конститутивный гетерохроматин. Возможно, это связано с тем, что в клетках таких мух мало белков, необходимых для образования гетерохроматина. Аналогичным действием обладают и многие генные мутации. Поскольку соответствующие гены клонированы и секвенированы, можно надеяться, что в скором будущем белки, участвующие в образовании гетерохроматина у дрозофилы, тоже будут известны.
Независимо от того, каковы молекулярные механизмы упаковки определенных областей генома эукариот в гетерохроматин, сам феномен гетерохроматизации следует отнести к таким регуляторным процессам, которые отличают клетки эукариот от клеток бактерий. Особенность такой уникальной формы регуляции состоит в том, что в данном случае память о функциональном статусе гена хранится в виде наследуемой структуры хроматина и не обусловлена существованием стабильной обратной связи саморегулирующихся белков-регуляторов, которые в ядре могут менять свою локализацию. Неизвестно, действуют ли механизмы такого типа лишь в случае инактивации больших областей хромосомы или же они могут работать и на уровне одного или нескольких генов. Данные, приведенные ниже, позволяют предположить, что экспрессия отдельных генов часто регулируется близлежащей контролирующей последовательностью и не зависит полностью от общего хромосомного окружения.

Рис. 10-38. Эффект положения мозаичного типа у дрозофилы. Распространению гетерохроматина (обозначен серым цветом) в соседние эухроматиновые области обычно препятствуют специальные пограничные последовательности неизвестной породы. Однако у мух, несущих в хромосомах определенные транслокации, эти пограничные последовательности отсутствуют. А. На ранних стадиях развития таких мух гетерохроматин начинает распространяться в соседнюю область хромосомы, продвигаясь в разных клетках на разные расстояния. Б. Такое распространение вскоре останавливается, однако достигнутый уровень распространения гетерохроматина наследуется; в результате получаются большие клоны клеток-потомков, у которых одни и те же гены сконденсированы в гетерохроматин и, следовательно, инактивированы (отсюда и «мозаичный» вид у некоторых из этих мух). Это явление имеет много общего с инактивацией Х-хромосомы у млекопитающих.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
10.3.11. Для оптимальной экспрессии гена часто бывает необходимо его определенное положение в хромосоме [30]
Гены, перенесенные в другие участки хромосомы, транскрибируются в зависимости от типа клетки. Следовательно, они должны нести с собой информацию, необходимую для их селективной экспрессии в соответствующих клетках. В качестве примера можно привести ген дрозофилы Sgs-З, которому для правильной транскрипции в слюнных железах необходимо 600 нуклеотидных пар, расположенных перед ним; установлено, что его транскрипция осуществляется с нормальной скоростью почти в любом сайте (но не в гетерохроматине) политенной хромосомы. Подобным образом трансгенные млекопитающие, содержащие фрагмент тканеспецифического гена, экспрессируют этот ген в соответствующих клетках, но только в том случае, если фрагмент достаточно длинен и несет большую часть последовательности энхансера.
Большинство привнесенных генов в клетках трансгенных животных для правильной экспрессии требуют гораздо больше фланкирующей последовательности, чем ген Sgs-З. Более того, у разных трансгенных особей наблюдаются разные уровни активности этих генов. Так как подобные вариации зависят от того, где трансген встроился в хромосому животного-хозяина, их называют эффектом положения в хромосоме. Некоторые результаты, полученные при встраивании гена альфа-фетопротеина мыши в случайные области ее генома, представлены в табл. 10-2. В данном примере ген активен только в тех тканях, где он экспрессируется и в норме, однако уровень его транскрипции обычно в пять-десять раз ниже, чем он должен быть в тех тканях, где его активность высока. И наоборот, уровень транскрипции может быть аномально высок в тканях, где обычно уровень экспрессии этого гена низок.
Если ген β-глобина человека (для которого характерен сильный эффект положения при экспрессии в эритроцитах трансгенных мышей)
соединить с фрагментом ДНК, который обычно расположен на расстоянии 50000 нуклеотидных пар от его промотора, эффект положения исчезает и полностью восстанавливается транскрипционная активность.
| Таблица 10-2, При экспрессии генов, перенесенных в геном мыши, обычно проявляется эффект положения. У независимо полученных трансгенных особей уровень активности генов различается | |||||
| Процент от тотальной мРНК в клетках | |||||
| желточного мешка | печени | кишечник | мозга | Копийность гена на клетку | |
| Эндогенный ген | 20 | 0,1 | 0 | 2 | |
| Трансгенная особь 1 | 3,4 | 0,1 | |||
| Трансгенная особь 2 | 4,8 | 1,3 | |||
| Трансгенная особь 3 | 4,4 | 4,7 | |||
| Трансгенная особь 4 | 0,4 | 0,4 |
В состав фрагмента ДНК, который был инъецирован в оплодотворенную яйцеклетку мыши, помимо гена альфа-протеина входила последовательность размером 14000 нуклеотидных нар. Эта последовательность содержала три энхансера, влияющих на экспрессию гена альфа-фетопротеина.
Сравнение уровней синтеза мРНК инъецированного гена и мРНК, образующейся в норме эндогенным геном в указанных тканях зародыша, проводили методом гибридизации. (По R. Е. Hammer et al., Science 235:53-58, 1987.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
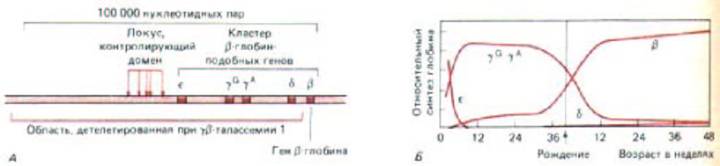
Рис. 10-39. Кластер β-подобных глобиновых генов у человека. А. Организация этих генов. Представлен большой участок хромосомы, содержащей 100000 нуклеотидных пар. Он содержит кластер сверхчувствительных к нуклеазе сайтов, составляющих «доменкон-тролирующий участок». Этот контролирующий локус и все, за исключением одного, глобиновые гены в случае γ- β -талассемии несут делецию. Б. Изменения при экспрессии β -подобных глобиновых генов на разных стадиях развития человека. Каждая из глобиновых цепей, контролируемых этими генами, для образования гемоглобина эритроцитов соединяется с α-глобиновой цепью, У пациентов с талас-семией уровень синтеза β -глобина в значительной степени снижен (не показано). (А-по F. Grosveld, G. В. van Assendelft, D. R. Greaves and G. Kollias, Cell 51: 975-985, 1987.)
Этот фрагмент, содержащий шесть сверхчувствительных к нуклеазе сайтов (см. разд. 9,1.19), оказывает воздействие на весь кластер глобиновых генов; его называют участком, контролирующим домен. Главный вывод, который следует из результатов изучения эффекта положения, состоит в том, что многие гены позвоночных для достижения нужного уровня экспрессии нуждаются в определенных последовательностях, расположенных на некотором расстоянии от них. Далее мы обсудим, какова роль этих последовательностей.
10.3.12. Для активации эукариотических генов может быть необходима локальная деконденсация хроматина [31]
У людей с определенной формой талассемии (наследственная форма анемии) перед геном β-глобина имеются большие делении ДНК.
Делетированная область (~ 100 000 нуклеотидных пар) содержит несколько β-глобин-подобных генов, а также участок, контролирующий домен, который был идентифицирован в экспериментах с трансгеннымя мышами (рис. 10-39, А). Хотя сам ген β-глобина и не поврежден, уровень его транскрипции значительно снижен. При обработке нуклеазой этот ген, в отличие от нормального гена β -глобина, демонстрирует такую же низкую
скорость реакции, что и основная фракция хроматина и, следовательно, не имеет структуры активного хроматина. Его нормальный гомолог в том же эритроците не содержит делеции, и к моменту начала транскрипции первого из этой группы гена (ген ε-глобина) весь кластер β-глобин-
подобных генов (90000 нуклеотидных пар), по-видимому, деконденсируется, превращаясь в активный хроматин (рис. 10-39,£).
Подобные результаты свидетельствуют в пользу двухступенчатой схемы индукции транскрипции генов высших эукариот. На стадии 1
весь хроматин в области, содержащей десятки тысяч нуклеотидных пар, превращается в относительно деконденсированную «активную» форму (рис. 10-40). Эта стадия может запускаться определенным типом белка-регулятора, который вызывает структурное изменение в близлежащем хроматине. Такое изменение распространяется от домен-контролирующего участка через всю петлю этого домена хроматина. На стадии 2 белки-
регуляторы, которые действуют на энхансеры и лежащие перед промотором элементы, регулируют транскрипцию определенных генов, локализованных внутри области экспонированного активного хроматина. Благодаря такому местному контролю сперва в желточном мешке зародыша экспрессируется ген ε-глобина человека, затем в печени эмбриона экспрессируются два гена γ-глобина и, наконец, ко времени рождения включаются гены β-глобина (рис. 10-39,Б).
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 499; Нарушение авторских прав?; Мы поможем в написании вашей работы!