КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Образование митотического веретена в М-фазе клетки сопровождается разительными изменениями
|
|
|
|
13-29
13.5. Механика клеточного деления [35]
В этом последнем разделе мы будем говорить о событиях в фазе М - кульминации клеточного цикла. В этот сравнительно короткий период хромосомы конденсируются, а содержимое родительской клетки, удвоившееся благодаря синтетической активности в предшествующей интерфазе, распределяется между двумя дочерними клетками (рис. 13-42).
По-видимому, на молекулярном уровне фаза М инициируется каскадом фосфорилирования белков, запускаемым при появлении М-
стимулирующего фактора (MPF), и заканчивается при дефосфорилировании, которое возвращает белки в их интерфазное состояние (разд. 13.2.5). В свою очередь фосфорилирование белков в течение М-фазы, вероятно, ответственно за многие морфологические изменения, сопровождающие митоз, в том числе и за конденсацию хромосом, разрушение ядерной оболочки и изменения цитоскелета, описанные ниже. Первое хорошо видимое проявление наступающей фазы М состоит в постепенном уплотнении дисперсного интерфазного хроматина в нитевидные хромосомы. Эта конденсация хромосом необходима для их последующего упорядоченного расхождения в дочерние клетки и сопровождается фосфорилированием многочисленных молекул гистона Н1, имеющихся в клетке (до шести фосфатных групп на одну молекулу HI). Поскольку гистон HI присутствует в количестве примерно одной молекулы на нуклеосому и известно, что он участвует в упаковке нуклеосом (разд. 13.2.5), то его фосфорилирование киназой MPF (разд. 9.1.12) в начале фазы М должно быть главной причиной конденсации хромосом. Такое молекулярное объяснение, пока еще гипотетическое, показывает, на каком уровне в конечном счете должен описываться весь клеточный цикл.
|
|
|
Кто-то сказал, что хромосомы в митозе напоминают покойника на похоронах: они дают повод для действий, но не принимают в них активного участия. Активная роль принадлежит двум особым цитоскелетным структурам, которые временно образуются в М-фазе. Первым появляется двухполюсное митотическое веретено, состоящее из микротрубочек и связанных с ними белков. Сначала оно выстраивает реплицированные хромосомы в плоскости деления клетки; затем каждая хромосома разделяется на две дочерние, которые разводятся нитями веретена к противоположным сторонам клетки. Вторая цитоскелетная структура, необходимая в М-фазе животных клеток, - это сократимое
кольцо из актиновых и миозиновых филаментов, появляющееся чуть позже под плазматической мембраной. Это кольцо втягивает мембрану внутрь, разделяя клетку на две, и тем самым обеспечивает, что каждая дочерняя клетка получит не только один полный набор хромосом, но и половину содержимого цитоплазмы и органелл родительской клетки. Эти две цитоскелетные структуры содержат разные наборы белков и в некоторых специализированных клетках могут формироваться независимо друг от друга. Однако их образование обычно тесно скоординировано,
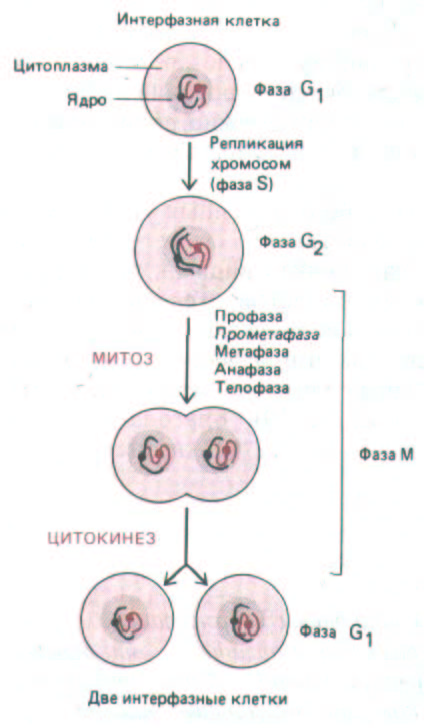
Рис. 13-42. М-фаза клеточного цикла начинается после фазы G2 и заканчивается к началу фазы G1 следующего цикла. Она состоит из пяти стадий деления ядра (митоза) и деления цитоплазмы (цитокинеза).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
так что разделение цитоплазмы (цитокинез) происходит сразу же после окончания деления ядра (митоза)', последнее относится и к растительным клеткам, хотя, как мы увидим, их жесткие стенки требуют иного механизма цитокинеза.
В сделанном только что описании речь шла об зукариотических клетках. Клетки бактерий не содержат ни актиновых филаментов, ни микротрубочек; в них обычно имеется только одна хромосома, и после ее репликации две копии распределяются между дочерними клетками с помощью механизма, который связан с прикреплением хромосомы к плазматической мембране бактерии (см. разд. 13.5.18). Сложный митотический аппарат, вероятно, стал необходим лишь с появлением клеток, содержавших гораздо большее количество ДНК в нескольких отдельных хромосомах. Главное назначение этого аппарата - точно распределять реплицированные хромосомы между двумя дочерними клетками.
|
|
|
Точность такого распределения была исследована на дрожжевых клетках, и оказалось, что одна ошибка приходится примерно на 105 клеточных делений.
13.5.1. М-фазу традиционно подразделяют на шесть стадий [35]
Основная стратегия деления клеток у зукариотических организмов удивительно постоянна. Первые пять стадий фазы М составляет митоз, шестой является цитокинез. Эти шесть стадий образуют динамическую последовательность, сложность и красоту которой трудно оценить по описаниям или по серии статических изображений. Описание митоза основано на наблюдениях двоякого рода: на результатах световой микроскопии живой клетки (нередко в сочетании с микрокиносъемкой) и на данных световой и электронной микроскопии фиксированных и окрашенных клеток. Различные стадии клеточного деления кратко описаны на схеме 13-1. Пять стадий митоза - профаза, прометафаза, метафаза, анафаза и телофаза - осуществляются в строго определенном порядке; цитокинез начинается во время анафазы и продолжается до конца митотического цикла (рис. 13-43). Световые микрофотографии деления типичной животной и типичной растительной клеток приведены на рис. 13-
44 и 13-45 соответственно.
В животном и растительном царствах встречаются бесчисленные вариации всех стадий деления, обобщенно представленных на схеме 13-
1. Мы будем упоминать некоторые из этих вариаций при более близком ознакомлении с механизмами деления клетки, так как они могут помочь нам понять действие разных частей митотического аппарата млекопитающих.
динамических свойств микротрубочек [36]
Из гл. 11 мы знаем, что главным центром организации микротрубочек у большинства животных клеток служит центросома - скопление аморфного материала, окружающее пару центриолей (разд. 11.4.4). Во время интерфазы материал центросомы инициирует рост микротрубочек, который направлен к периметру клетки, в то время как их начальные участки (минус-концы) остаются связанными с центросомой. Это интерфазное скопление микротрубочек, расходящихся от центросомы, представляет собой динамичную, непрерывно меняющуюся структуру, в которой отдельные микротрубочки все время возникают и распадаются. Новые микротрубочки растут путем пристраивания молекул тубулина к плюс-концам; спорадически и, по-видимому, случайно индивидуальные микротрубочки становятся нестабильными и подвергаются быст-
|
|
|
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
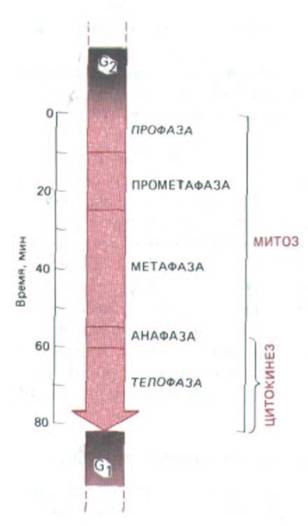
Рис. 13-43. Временной ход митоза и цитокинеза, типичный для клетки млекопитающего. Точные цифры для разных клеток различны.
Обратите внимание, что цитокинез начинается еще до окончания митоза. Началом профазы (и, следовательно, фазы М в целом) считают тот момент клеточного цикла, когда впервые становятся видимыми конденсированные хромосомы; это несколько произвольный критерий, так как степень конденсации хромосом постепенно увеличивается уже в поздней фазе G2.
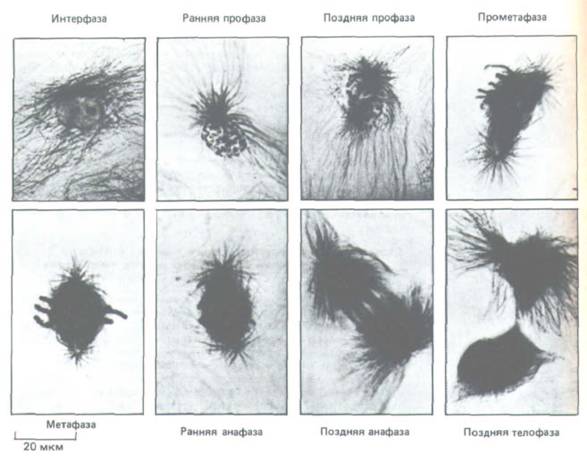
Рис. 13-44. На этих световых микрофотографиях культивируемых клеток сумчатого (клеток Р.К) показан ход митоза в животной клетке.
Микротрубочки видны благодаря окрашиванию антителами с золотом; хроматин окрашен толуидиновым синим. Главные события митоза на уровне световой микроскопии известны уже более 100 лет. В интерфазе центросома, содержащая пару центриолей, служит центром интерфазного скопления микротрубочек. В ранней профазе единственная центросома содержит две пары центриолей (на снимке не видны); в поздней профазе
центросома делится, в результате чего образовавшиеся звезды отходят друг от друга. В прометафазе разрушается ядерная оболочка, и это позволяет микротрубочкам веретена взаимодействовать с хромосомами. В метафазе уже ясно видна двухполюсная структура веретена и все хромосомы выстраиваются в его экваториальной области. В ранней анафазе все хроматиды одновременно разделяются и под действием нитей веретена начинают двигаться к полюсам. В течение поздней анафазы полюса веретена все дальше отходят друг от друга, еще более раздвигая две группы хроматид. В телофазе формируются дочерние ядра, и в поздней телофазе почти полностью завершается цитокинез; между дочерними клетками сохраняется остаточное тельце. (Фотографии любезно предоставлены М. de Brabander.)
|
|
|
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
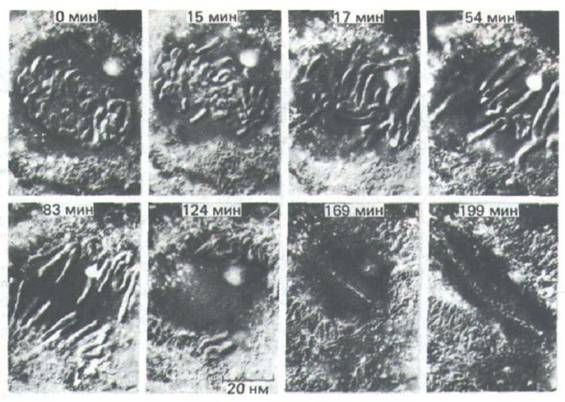
Рис. 13-45. Ход митоза в типичной растительной клетке. Микрофотографии живой клетки Haemanthus (лилейные), полученные с помощью метода дифференциального интерференционного контраста (разд. 4.1.5). Необычно крупные хромосомы в этой клетке легко наблюдать.
А. Профаза: хромосомы сконденсировались и ясно видны в ядре клетки (N). Б и В. Прометафаза: ядерная оболочка разрушена и хромосомы взаимодействуют с микротрубочками, отходящими от двух полюсов веретена (Р). Обратите внимание на то, что между представленными здесь стадиями (Б и В) прошло только две минуты. Г. Метафаза: хромосомы расположились в виде метафазной пластинки, а их кинетохоры находятся посередине между обоими полюсами веретена. Д. Анафаза: хромосомы разделились на сестринские хроматиды, которые теперь движутся к противоположным полюсам. Е. Телофаза: хромосомы деконденсируются, образуя два ядра, которые будут видны позже. Ж и 3. Цитокинез:
показаны две последовательные стадии формирования клеточной пластинки; она видна как линия, направления роста которой указаны стрелками (3). (С любезного разрешения Andrew Bajer.)
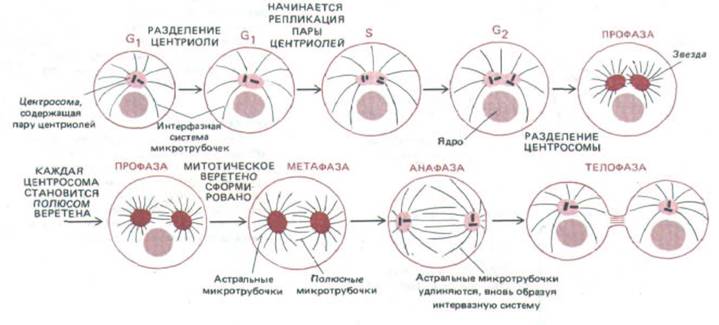
Рис. 13-46. Центросомный цикл. В интерфазной клетке центросома удваивается с образованием двух полюсов митотического веретена. В большинстве животных (но не растительных) клеток пара центриолей (показанных как пара коротких черных отрезков) погружена в материал центросомы (выделен цветом), от которого растут микротрубочки. В определенный момент фазы Gt две центриоли расходятся на несколько микрон. В течение фазы S возле каждой старой центриоли под прямым углом к ней начинает формироваться дочерняя центриоль. Рост дочерних центриолей обычно завершается в фазе G2. Вначале обе пары центриолей остаются погруженными в единую массу центросомного материала, образующего одну центросому. В ранней фазе М каждая пара центриолей становится частью отдельного центра организации микротрубочек, от которого отходит радиальный пучок микротрубочек - звезда. Две звезды, первоначально лежавшие бок о бок около ядерной оболочки, теперь отходят друг от друга. В поздней профазе пучки полюсных микротрубочек, принадлежащие двум звездам и взаимодействующие между собой, избирательно удлиняются, по мере того как два центра расходятся по двум сторонам ядра. Таким способом быстро формируется митотическое веретено.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
Схема 13-1. Шесть стадий клеточного деления.
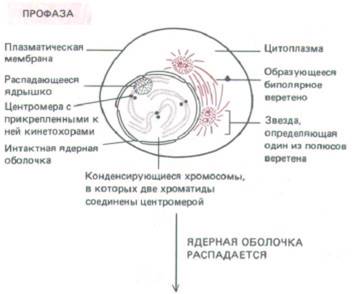
| ПРОФАЗА Переход из фазы G2 в фазу М, как это видно в микроскоп, совершается постепенно. Хроматин, который в интерфазе выглядит диффузным, конденсируется в отчетливо видимые хромосомы. Для каждого вида характерно совершенно определенное число хромосом. Каждая хромосома во время предшествующей фазы S радуплицировалась и состоит теперь из двух сестринских хроматид. В каждой из хроматид имеется специфический участок ДНК, называемый центромерой, который необходим для их правильного расхождения. В конце профазы цитоплазматические микротрубочки, составляющие часть интерфазного цитоскелета, распадаются и начинается образование веретена — главного компонента митотического аппарата. Веретено представляет собой двухполюсную структуру, состоящую из микротрубочек и связанных с ними белков. Сборка веретена происходит вначале вне ядра. |
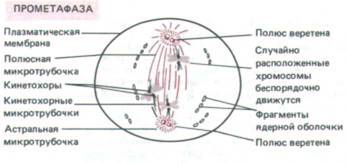
| ПРОМЕТАФАЗА Про метафаза начинается с быстрого распада ядерной оболочки на мелкие мембранные пузырьки, неотличимые от фрагментов эндоплазматического ретикулума. Эти пузырьки остаются видимыми около веретена во время митоза. Микротрубочки веретена, которые находились вне ядра, могут теперь проникнуть в ядерную область. У хромосом на каждой центромере образуются особые белковые комплексы, называемые кинетохорами; они прикрепляются к некоторым из микротрубочек веретена, получающим теперь название кинетохорных микротрубочек. Остальные микротрубочки веретена называют полюсными, а те, которые лежат вне веретена, — астральными. Кинетохорные микротрубочки идут в противоположных направлениях от двух сестринских хроматид каждой хромосомы и тянут их в разные стороны, что приводит к интенсивному движению хромосом. |
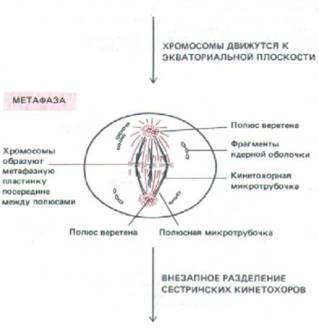
| МЕТАФАЗА Кинетохорные микротрубочки в конце концов приводят каждую хромосому в экваториальную плоскость на полпути между полюсами веретена. Хромосомы образуют здесь метафазную пластинку, в которой они удерживаются натяжением кинетохорных микротрубочек, отходящих от них к противоположным полюсам веретена. |
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
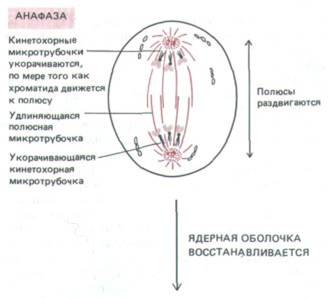
| АНАФАЗА Запускаемая специфическим сигналом, анафаза начинается с внезапного разделения парных кинетохоров каждой хромосомы, после чего ее две хроматиды начинают медленно расходиться к соответствующим полюсам. Все хроматиды движутся с одинаковой скоростью около 1 мкм/мин. Здесь можно различать движение двоякого рода. Во время анафазы А кинетохорные микротрубочки укорачиваются, по мере того как хромосомы приближаются к полюсам. Во время анафазы В происходит удлинение полярных микротрубочек и полюсы веретена еще дальше отодвигаются друг от друга. Анафаза обычно длится всего лишь несколько минут. |
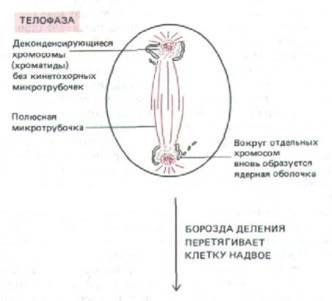
| ТЕЛОФАЗА В телофазе (от греч. telos — конец) разделившиеся дочерние хроматиды подходят к полюсам и кинетохорные микротрубочки исчезают. Полярные микротрубочки продолжают удлиняться, после чего вокруг каждой группы дочерних хроматид образуется новая ядерная оболочка. Конденсированный хроматин начинает разрыхляться, появляются вновь ядрышки (исчезнувшие в профазе), и митоз заканчивается. |

| ЦИТОКИНЕЗ Это процесс разделения цитоплазмы, он обычно начинается где-то в анафазе. На схеме показано, как он протекает в животных клетках. Мембрана в экваториальной области (между двумя дочерними ядрами) начинает втягиваться внутрь по направлению к оси веретена; в результате образуется борозда деления, которая постепенно углубляется, пока не дойдет до остатков веретена, расположенного между ядрами. Этот мостик, называемый остаточным тельцем, может некоторое время сохраняться, а затем разрушается, что ведет к полному разделению дочерних клеток. |
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
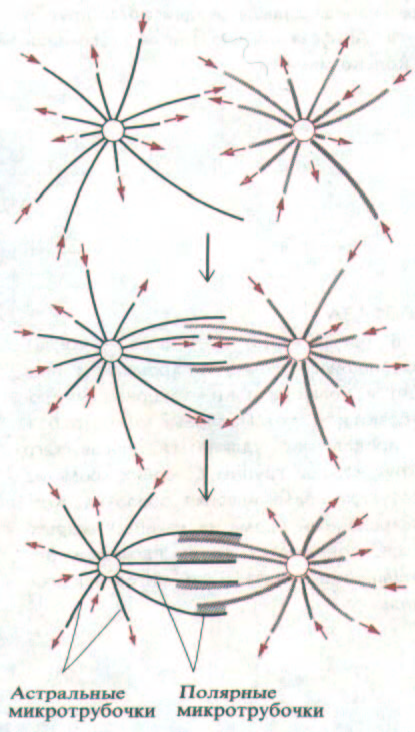
Рис. 13-47. Модель образования двухполюсного) митотического веретена путем селективной стабилизации взаимодействующих микротрубочек.
Новые микротрубочки отрастают в случайных направлениях от двух центросом (представленных кружками), к которым они прикреплены своими минус-концами. Их плюс-концы «динамически нестабильны» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка (разд. 11.4.3). Когда две микротрубочки от противоположных центросом взаимодействуют в зоне их перекрывания, белки, связанные с микротрубочками, сшивают их друг с другом (показано серым цветом), прикрывая и стабилизируя таким образом их плюс-концы и уменьшая вероятность деполимеризации.
рому, «катастрофическому» распаду, добавляя свои молекулы-субъединицы к пулу неполимеризованного тубулина, содержащегося в цитоплазме (см. схему 11-2). I Как показано на рис, 13-46, в центросоме на протяжении всего1 клеточного цикла происходят изменения. Где-то в фазе S пара центриолей реплицируется, оставаясь внутри одного скопления центросомного материала. В профазе центросома расщепляется и каждая дочерняя центросома становится центром отдельной звезды - структуры из микротрубочек, концы которых погружены в материал центросоми. Микротрубочки обеих звезд удлиняются до соприкосновения друг с другом, после чего две центросомы расходятся. Затем, в прометафазе, ядерная оболочка разрушается, и это позволяет микротрубочкам от каждой центросомы проникать в ядро и взаимодействовать с хромосомами. Две дочерние центросомы называют теперь двумя полюсами веретена.
Как полагают, только что описанные события обусловлены происходящими в профазе существенными изменениями стабильности микротрубочек и свойств центросомы. Мы уже упоминали о том, что, судя по имеющимся данным, фактор MPF вызывает переход в фазу М, инициируя каскад фосфорилирования целого ряда белков (разд. 13.2.5).
При этом фосфорилируются некоторые молекулы, взаимодействующие с микротрубочками, поскольку при переходе клетки в профазу время полужизни средней микротрубочки уменьшается примерно в 20 раз (от примерно 5 мин до 15 с, см. рис. 13-48). Это, видимо, связано с резким повышением вероятности того, что типичная растущая микротрубочка начнет укорачиваться в результате какого-то изменения на ее плюс-
конце (разд. 11.4.3, схема 11-2), а также происходящего в профазе изменения центросомы, сильно увеличивающего ее способность к образованию новых микротрубочек (это можно наблюдать in vitro). Этих двух изменений достаточно для того, чтобы объяснить, почему в начале фазы М отмечается быстрый переход от сравнительно малого числа длинных микротрубочек, отходящих от центросомы к периферии клетки (интерфазное скопление микротрубочек), к большому числу коротких микротрубочек, окружающих каждую центросому (см. профазу на рис. 13-46).
Полагают, что в ходе митоза удлиняющиеся концы микротрубочек, отходящих от полюсов веретена, наталкиваются на структуры, которые связываются с ними и стабилизируют их против катастрофического разрушения. Поскольку каждый полюс испускает микротрубочки в разных направлениях, такая избирательная стабилизация и создает характерную двухполюсную форму митотического веретена, в котором большинство микротрубочек отходит от двух полюсов к экваториальной пластинке, находящейся на полпути между ними. Те микротрубочки, которые пересекают экватор, могут селективно стабилизироваться присоединяющимися к ним белками - эти белки сшивают соседние параллельные микротрубочки противоположной полярности (рис. 13-47). В метафазе веретено в клетках высших животных и растений может содержать до нескольких тысяч микротрубочек, тогда как у некоторых грибов их всего лишь около 40.
Хотя некоторые микротрубочки в веретене частично стабилизированы против спонтанного разрушения, большинство из них продолжает обмениваться своими субъединицами с пулом растворенных молекул тубулина в цитозоле. Этот обмен может быть непосредственно измерен с помощью метода, представленного на рис. 13-48. Его можно также выявить, помещая митотические клетки в условия, обратимо сдвигающие равновесие между полимеризацией и деполимеризацией тубулина, и наблюдая двойное лучепреломление микротрубочек веретена в поляризованном свете (рис. 13-49). Если митотические клетки поместить
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
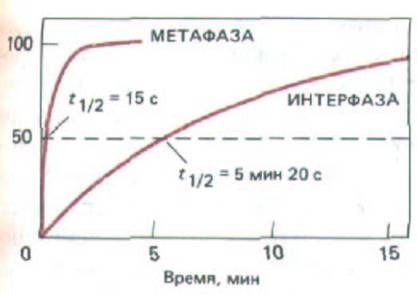
Рис. 13-48. Результаты исследования, показывающие, что микротрубочки в М-фазной клетке в среднем гораздо более динамичны, чем в интерфазе. В клетки млекопитающих в культуре инъецировали тубулин с ковалентне присоединенным флуоресцентным красителем. По прошествии времени, необходимото для включения флуоресцентного тубулина в микротрубочки, в небольшом участке весь тубулин обесцвечивали интенсивным лазерным лучом. Восстановление флуоресценции в облученном участке микротрубочек, вызванное заменой их обесцвеченных субъединиц необесцвеченным флуоресцентным тубулином из пула субъединиц в растворе, было затем представлено в виде функции времени.
Время 50%-ного восстановления флуоресценции, ti,2, как полагают, равно времени, необходимому для того, чтобы половина микротрубочек в этом участке деполимеризовалась и восстановилась. (По данным W. М. Saxon et al., J. Cell Biol. 99: 2175-2187, 1984, by copyright permission of the Rockefeller Univ. Press.)
в тяжелую воду (D2O) или обработать таксолом (эти воздействия подавляют разборку микротрубочек), то нити веретена будут удлиняться. Такое стабилизированное веретено не может тянуть хромосомы, и митоз останавливается. Но митоз блокируется и при прямо противоположном воздействии, если нити веретена обратимо разрушить с помощью одного из трех агентов, подавляющих сборку тубулина в микротрубочки, -
колхицина, низкой температуры или высокого гидростатического давления. Тот факт, что ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы, указывает на то, что для правильного функционирования веретена необходимо тонкое равновесие между сборкой и разборкой. Прежде чем рассматривать механизм таких движении, мы опишем более подробно организацию веретена и расположение хромосом в митозе.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2471; Нарушение авторских прав?; Мы поможем в написании вашей работы!