КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Основные этапы холинергической передачи и их фармакологическая коррекция
|
|
|
|
1. Синтез и депонирование медиатора. Ацетилхолин синтезируется в пресинаптических окончаниях из ацетил-КоА и холина. В цитоплазме пресинаптического окончания содержится большое количество митохондрий, здесь путем окислительного декарбоксилирования a-кетокислот синтезируется ацетил-КоА. Холин поступает в клетку извне благодаря специальному трансмембранному переносчику. Транспорт холина в нейрон сопряжен с переносом ионов натрия и может быть блокирован гемихолином.
Таблица 2. Сравнительная характеристика холинорецепторов клетки.
| Тип | Агонист | Антагонист | Локализация | Функция | Механизм |
| НМ | PTMA Никотин | d-тубокурарин a-бунгаротоксин | Скелетные мышцы | Деполяризация концевой пластинки, сокращение мышцы | Открытие Na+-канала |
| НН | DMPP Эпибатидин Никотин | Триметафан | Вегетативные ганглии Мозговое вещество надпочечников Каротидные клубочки ЦНС | Деполяризация и возбуждение постганглионарного нейрона Секреция адреналина и норадреналина Рефлекторная стимуляция дыхательного центра Контроль психических и моторных функций, когнитивные процессы. | Открытие Na+, K+ и Са2+-каналов |
| М1 | Мускарин Оксотреморин | Атропин Пирензепин | Вегетативные ганглии (пресинаптически) ЦНС | Деполяризация, усиление секреции медиатора (поздний постсинаптический потенциал) Контроль психических и моторных функций, когнитивные процессы. | Активация фосфолипазы С через Gq белок и синтез IP3 (выход Са2+ из депо), DAG (активация Са2+-каналов, протеинкиназы С). |
| М2 | Мускарин Метахолин | Атропин Метоктрамин Трипитрамин | Миокард | САУ: снижение автоматизма; АВУ: снижение проводимости; Рабочий миокард: незначительное снижение сократимости. | Через a-единицу Gi-белка торможение аденилатциклазы (”цАМФ). Через bg-единицы Gi-белка активация К+-каналов и блокада L-типа Са2+-каналов. |
| М3 | Бетанехол | Атропин Дарифенацин HHSDP | Гладкие мышцы Железы Эндотелий сосудов (внесинаптически) | Сокращение, “ тонуса Повышение секреции Секреция NO и дилятация сосуда | Подобен М1 |
| М4 | ? | ? | Сердце Альвеолы ЦНС | ? | Подобен М2 |
| М5 | ? | ? | Слюнные железы Радужка глаза Моноциты ЦНС | ? | Подобен М1 |
Примечание: a-бунгаротоксин – яд тайваньской гадюки Bungaris multicintus и кобры Naja naja.
|
|
|
PTMA – фенилтриметиламмоний
DMPP – диметилфенилпиперазин
HHSDP – гексагидросиладифенол
АВУ – атриовентрикулярный узел
САУ – синоаурикулярный узел
Синтез ацетилхолина проводит особый фермент холинацетилтрансфераза, путем ацетилирования холина. Образовавшийся ацетилхолин поступает в везикулы при помощи антипортера переносчика в обмен на протон. Работа этого переносчика может быть заблокирована векзамиколом. Обычно в каждой везикуле содержится от 1.000 до 50.000 молеукл ацетилхолина, а общее число везикул в пресинаптическом окончании достигает 300.000.
2. Выделение медиатора. Во время фазы покоя, через пресинаптическую мембрану выделяются единичные кванты медиатора (изливается содержимое 1 везикулы). Одна молекула ацетилхолина способна вызвать изменение потенциала мембраны всего на 0,0003 мВ, а то количество, которое содержится в 1 везикуле – на 0,3-3,0 мВ. Такие миниатюрные сдвиги не вызывают развития биологического ответа, но поддерживают физиологическую реактивность, тонус ткани-мишени.
Активация синапса происходит в тот момент, когда на пресинаптическую мембрану приходит потенциал действия. Под влиянием потенциала мембрана деполяризуется и это вызывает открытие воротного механизма медленных кальциевых каналов. По этим каналам ионы Са2+ поступают в пресинаптическое окончание и взаимодействуют с особым белком в мембране везикул – синаптобревином (VAMP). Синаптобревин переходит в активированное состояние и начинает выполнять роль своеобразного «крючка» или якоря. Этим якорем везикулы фиксируются к пресинаптической мембране в тех местах, где лежат особые белки – SNAP-25 и синтаксин-1. В последующем эти белки инициируют слияние мембраны везикул с мембраной аксона и выталкивают медиатор в синаптическую щель подобно поршню насоса. При прохождении потенциала действия через пресинаптическую мембрану одновременно опустошаются 2.000-3.000 везикул.
|
|
|
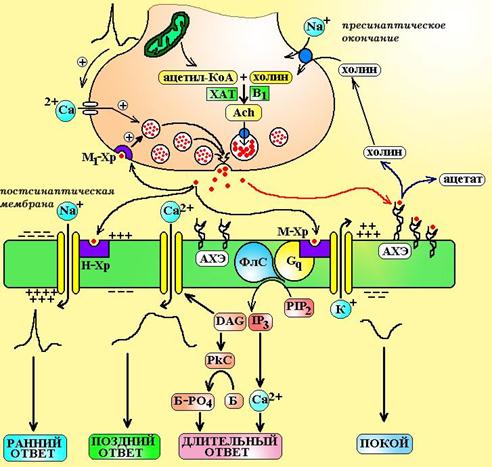
Схема 4. Передача сигнала в холинергическом синапсе. ХАТ – холинацетилтрансфераза, В1 – тиамин, Ach – ацетилхолин, М1-Хр – М1-холинорецепторы, АХЭ – ацетилхолинэстераза, ФлС – фосфолипаза С, PIP2 – фосфатидилинозитол бифосфат, IP3 – инозитол трифосфат, DAG – диацилглицерол, PkC – протеинкиназа С, Б – белок-фермент, Б-РО4 – фосфорилированная форма белка-фермента.
Процесс выделения медиатора может быть нарушен под влиянием ботулотоксина (токсин бактерий Clostridium botulinum). Ботулотоксин вызывает протеолиз белков, участвующих в выделении медиатора (SNAP-25, синтаксин, синаптобревин). a-латротоксин – яд паука «черная вдова» связывается с белком SNAP-25 (нейрексином) и вызывает спонтанный массивный экзоцитоз ацетилхолина.
3. Развитие биологического ответа. В синаптической щели путем диффузии ацетилхолин поступает к постсинаптической мембране, где активирует холинорецепторы. При взаимодействии с Н-холинорецепторами происходит открытие натриевых каналов и на постсинаптической мембране генерируется потенциал действия.
В том случае, если ацетилхолин активирует М-холинорецепторы, сигнал передается через систему G-белков на фосфолипазу С, ионные каналы К+ и Са2+ и все это приводит в конечном итоге к изменению поляризации мембраны, процессов фосфорилирования внутриклеточных белков.
Помимо постсинаптической мембраны ацетилхолин может воздействовать на холинорецепторы пресинаптической мембраны (М1 и М2). При активации ацетилхолином М1-пресинаптического рецептора выделение медиатора усиливается (положительная обратная связь). Роль М2-холинорецепторов на пресинапетической мембране недостаточно ясна, полагают, что они могут тормозить секрецию медиатора.
|
|
|
Развитие биологического ответа можно вызвать при введении лекарственных веществ, которые стимулируют холинорецепторы или предотвратить, если ввести средства, блокирующие эти рецептры. Повлиять на развитие эффекта можно и не затрагивая рецепторы, а воздействуя лишь на пострецепторные механизмы:
· Токсин коклюшной палочки может активировать Gi-белок и снижать активность аденилатциклазы на затрагивая М-холинорецептор;
· Токсин холерного вибриона может активировать Gs-белок и повышать активность аденилатциклазы;
· Дитерпен форсколин из растения Coleus forskohlii способен непосредственно активировать аденилатциклазу в обход рецепторов и G-белков.
4. Окончание действия медиатора. Время существования ацетилхолина в синаптической щели составляет всего 1 мС, после чего он подвергается гидролизу до холина и остатка уксусной кислоты. Уксусная кислота быстро утилизируется в цикле Кребса. Холин в 1.000-10.000 раз менее активен, чем ацетилхолин, 50% его молекул подвергаются обратному захвату в аксон для ресинтеза ацетилхолина, остальная часть молекул включается в состав фосфолипидов.
Гидролиз ацетилхолина осуществляет особый фермент – холинэстераза. В настоящее время известно 2 его изоформы:
· Ацетилхолинэстераза (AChE) или истинная холинэстераза – осуществляет высокоспецифичный гидролиз ацетилхолина и локализуется на постсинаптической мембране холинергических синапсов.
· Бутирилхолинэстераза (ButChE) или псевдохолинэстераза – осуществляет низкоспецифичный гидролиз эфиров. Локализуется в плазме крови и перисинаптическом пространстве.
Сравнительная характеристика этих ферментов представлена в таблице 3.
Таблица 3. Сравнительная характеристика холинэстераз.
| Параметр | Ацетилхолинэстераза | Бутирилхолинэстераза |
| Источник Распространение | Холинергические нейроны Все холинергические нейроны, эритроциты, серое вещество мозга | Гепатоциты Плазма, печень, кишечник, белое вещество |
| Субстраты гидролиза · Ацетилхолин · Метахолин · Бутирилхолин | Очень быстро Быстро Не гидролизуется | Медленно Не гидролизуется Медленно |
| Антагонисты | Более чувствительна к физостигмину | Более чувствительна к ФОС |
| Функция | Окончание действия ацетилхолина | Гидролиз эфиров пищи |
|
|
|
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 412; Нарушение авторских прав?; Мы поможем в написании вашей работы!