КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Тема 3: геномы эукариот
|
|
|
|
Схема фиксации лидерной последовательности И-РНК на комплементарной последовательности 16S РНК рибосомы
IK – инициирующий кодон АТГ, который кодирует на РНК – АУГ, с него при трансляции начинается синтез полипептида. АУГ в составе молекулы и-РНК на рибосомах кодирует первую аминокислоту белка формилметионин (у эукариот – метионин). У некоторых микроорганизмов вместо АУГ может быть ГУГ, кодирующий валин.
Зона структурных генов(1-3) начинается сразу за инициирующим кодоном и содержит нуклеотидные последовательности генов, кодирующих ферменты катаболизма лактозы. Для того, чтобы белковые продукты этих генов в процессе трансляции на рибосомах могли разделяться на отдельные белковые молекулы ферментов, между структурными генами моут быть расположены дополнительные последовательности Шайна-Дальгарно.
Терминатор (терминаторный участок) начинается за зоной структурных генов. Он обеспечивает окончание процессов транскрипции (синтеза и-РНК) и трансляции на рибосомах. Терминатор состоит из трех важных блоков: ТК (терминирующего кодона), PD (ГЦ-палиндрома) и ТА-зоны. Рассмотрим их.
ТК -- терминирующий кодон важендля окончания трансляции на рибосомах.Может быть представлен такими триплетами на ДНК: ТАА, ТАГ или ТГА. На и-РНК – это будут стоп-кодоны или нонсенс-кодоны (бессмысленные) УАА, УАГ или УГА. Они не кодируют ни одной аминокислоты, поэтому при трансляции на рибосомах на них обрывается синтез полипептида.
PD -- палиндромы – зона терминатора. Это - инвертированные (повёрнутые на 180 градусов) последовательности, преимущественно из ГЦ- и ЦГ-пар, которые на ДНК способны образовывать крестовую, а на РНК – шпилечную структуру. Для палиндромов характерна комплементарность по горизонтали и вертикали. Именно ГЦ-палиндром в составе терминатора гена (оперона) приводит к замедлению продвижения по данному участку РНК-полимеразы, ведущей синтез и-РНК, т.е. тормозит транскрипцию.
|
|
|
ТА – зона терминатора – участок ДНК, в котором многократно повторяются ТА-пары нуклеотидов, т.е. в смысловой цепи ДНК повторяются остатки тимина, а в матричной - аденина. На матричной цепи ТА-участка ДНК при транскрипции РНК-полимераза синтезирует концевой участок информационной РНК – так называемый уридиловый хвост, состоящий из остатков урацила. Между адениловыми остатками ДНК-овой матрицы и комплементарными уридиловыми остатками синтезируемой и-РНК имеется только по две водородных связи, в отличие от ГЦ-пар, соединенных тремя водородными связями. Поэтому в ТА-зоне происходит легкое отсоединение и-РНК от ДНК-матрицы. Обычно ТА-зона терминатора достаточно протяженная в ρ -независимых (ро-независимых) терминаторах. В ρ -зависимых терминаторах основную роль в терминации (окончании) транскрипции выполняет ρ-белок. Этот белок «садится» на 5/-конец и-РНК, скользит по ней, догоняя РНК-полимеразу и после прокатки через ГЦ- палиндром, сбивает РНК-полимеразу с матричной цепи ДНК. Поэтому ТА зона в ρ -зависимых терминаторах может отсутствовать или быть слабо выражена.
Экспрессия генов у прокариот включает транскрипцию и трансляцию, однако из-за отсутствия ядерной мембраны эти процессы сопряжены и проходят почти одновременно. Освобождающаяся от матричной ДНК молекула и-РНК поступает на полисому (комплекс ≈ из 10 рибосом), которая располагается на ЦПМ, где почти сразу начинается трансляция. Белковый продукт появляется в клетке через 2,5-3 минуты после начала транскрипции.
Регуляция экспрессии генов осуществляется на уровне транскрипции и трансляции. На уровне транскрипции с помощью таких механизмов, как негативный и позитивный контроль, индукция и репрессия, аутогенный контроль, катаболитная репрессия. На уровне трансляции изучен механизм регулирования с помощью образования альтернативных шпилек на и-РНК - аттенуация.
|
|
|
In vivo (в живой клетке) чаще всего имеют место смешанные механизмы регулирования. Выделяют 4 типа классических оперонов:
1) индуцибельный с положительным контролем;
2) индуцибельный с отрицательным контролем;
3) репрессибельный с положительным контролем;
4) репрессибельный с отрицательным контролем;
Встречаются и смешанные механизмы регуляции, например, катаболитная репрессия.
Передача генетической информации у прокариот осуществляется как по вертикали (от материнской клетке к дочерней), так и по горизонтали (между клетками одного вида, разных видов, родов, семейств) с помощью механизмов рекомбинации – конъюгации, трансформации, трансдукции, сексдукции, трансфекции, транспозиции. Горизонтально, могут передаваться плазмиды, транспозоны, вирусы. Все они могут захватывать и переносить фрагменты генома.
Для прокариотов, кроме хромосомного, характерен еще плазмидный геном. В клетке может находиться от нескольких единиц до нескольких сотен плазмид.
Плазмиды – двухцепочечные кольцевые ДНК, которые несут дополнительные гены: гены устойчивости к токсинам, антибиотикам, сульфаниламидам, тяжёлым металлам, нефтепродуктам, нафталину, камфоре, а также гены синтеза токсинов, антибиотиков, гемолизинов и других факторов патогенности. Гены факторов патогенности обычно собраны в особые кластеры (группы), получившие название островов патогенности. С помощью плазмид, фагов и транспозонов острова патогенности и другие геномные острова (Genom islands) могут переноситься горизонтально от одной клетки к другой не только в пределах вида, но и между представителями разных родов и семейств.
В 70-х годах у бактерий обнаружили транспозоны – мобильные генетические элементы (так называемые «прыгающие» гены), способные перемещаться с одного сайта генома в другой, например, между различными участками хромосом, из хромосомы в плазмиду, в фаговую ДНК и наоборот.
Геном эукариот более сложен, чем у прокариот и включает нуклеотидные последовательности хромосом, ДНК митохондрий и пластид (1-10 % от общего генома, у дрожжей до 20%), ДНК плазмид у дрожжей, ДНК латентных и дефектных вирусов.
|
|
|
Ядро эукариот хорошо выражено, имеется ядерная мембрана, окружающая хромосомы. Хромосом много, они парные, состоят из гомологичных хроматид, каждая из которых представляет двухцепочечную молекулу ДНК (набор хромосом диплоидный). В составе хромосомы – 50 % ДНК и 50 % белков, которые представлены основными гистоновыми белками, входящими в состав нуклеосом, и кислыми белками, которые заполняют полость нуклеосом, разрыхляют ее и играют важную роль в распаде нуклеосом перед началом транскрипции и репликации.
В релаксированном состоянии хромосомы эукариот могут достигать нескольких сантиметров (у человека до 5 см в длину). Существуют несколько стадий конденсации хромосом, в результате чего хромосома компактизируется, накручивается на нуклеосомы и образует более сложные свернутые структуры.
Стадии компактизации (конденсации) хромосом. Акты конденсации и деконденсации хромосом сменяют друг друга в клеточном цикле: в интерфазе ДНК выглядит в виде вытянутых спутанных нитей и получила название - хроматин. В этом состоянии ДНК частично релаксирована, что облегчает прохождение процесса транскрипции и репликации. Для расхождения (сегрегации) хромосом в митозе очень важно, чтобы хромосомы были суперспирализованы – конденсированы. Для этого в начале профазы митоза ДНК начинает компактизоваться с помощью положительной и отрицательной суперспирализации, а также путем накручивания на нуклеосомы. Нуклеосомная нить ДНК напоминает бусы, в которых нить (суперспирализованная молекула ДНК) намотана на бусинки (нуклеосомы).

Рис. 3.1. Cтадии компактизации хроматина
Нуклеосома – октомер из 8 субъединиц белков-гистонов, включающая по 2 молекулы гистонов Н2А, Н2В, Н3, Н4. Диаметр нуклеосомы - 11 нм, высота - 5,7 нм. По краям от нуклеосом имеются свободные участки ДНК в 20-90 пар нуклеотидов – линкеры. Гистон Н1 не входит в состав нуклеосомы, а фиксирует петли линкеров, удерживая ДНК на нуклеосоме. Такое нуклеосомное строение хромосом характерно только для линейных хромосом эукариот.
|
|
|
В результате спирализации и накручивания на нуклеосомы хромосомы укорачиваются и превращаются в метафазные хромосомы (стадия метафазы), сокращаясь в длину в 10000 раз, а в диаметре – примерно в 700 раз. Это способствует нормальному расхождению (сегрегации) хромосом в анафазе митоза. Рентгеноструктурный анализ позволил выявить следующие стадии компактизации ДНК.
1-ая стадия - двухцепочечная спираль ДНК (диаметр – 2 нм), обычно в правозакрученной В-форме.
2-ая стадия – нуклеосомная нить (диаметр – 11нм). ДНК наматывается на частички нуклеосом, образуя на них 1,75 витка (146 пар нуклеотидов).
3-ая стадия – образование хроматиновой фибриллы (диаметр 30 нм). Нуклеосомы сближаются друг с другом, образуется зигзагообразная «лента», которая скручивается в соленоид – спираль с полостью внутри.
4-ая стадия – образование петлевых доменов (диаметр 300 нм) формируется путем формирования петель из нити соленоида.
5-ая стадия – образование метафазных хромосом, которые получили название «ламповых щеток» (диаметр 1400 нм).
Избыточность геномов эукариот. Только незначительная часть ДНК у эукариот представлена структурными и регуляторными генами, остальная часть генома представляет собой «эгоистичную» (сателлитную) ДНК, которая очевидно попала в геном эукариот путём интеграции вирусов и других мобильных генетических элементов. В геноме человека насчитывается 3,5 х 109 пар нуклеотидов. Геномы млекопитающих различаются, но имеют близкие значения молекулярной массы хромосом, достигающие сотен миллиардов Да. В соответствии с величиной генома у человека должно бы было быть 150000 и более генов, однако в 2003 г. американские ученые заявили о существовании 30000 генов, в последние годы предполагается наличие 75 тыс. генов, остальная часть геномной ДНК очевидно является “генетическим мусором”. Значительная часть генома представлена некодирующими последовательностями. У человека некодирующие последовательности составляют 80-85% (по другим данным – 92%), а у растений – до 90% и более, т.е. характерна избыточность генома.
В геноме эукариот выделяют следующие типы последовательностей ДНК:
1) многократно повторяющиеся последовательности, которых насчитывается более 105 повторов на геном. Чаще всего это блоки из 5-8 нуклеотидов, которые тандемно повторяются и образуют фрагменты в 150-500 пар нуклеотидов, например - (ААТАТ)30-100. Функция их до конца неизвестна, но предполагают, что они могут играть роль в регуляции работы генов – находятся в области центромер, теломер, интронов, транспозонов. Это последовательности: Alu, B1, B2, L1. Среди многократно повторяющихся последовательностей очень часто встречаются сайты рестрикции в составе палиндромов (см. дальше - тема «Репарации»). Сайты рестрикции могут быть теми горячими точками, куда встраиваются плазмиды, транспозоны, вирусные ДНК, трансгены.
2) умеренно повторяющиеся последовательности – встречаются на геном от 10 до105. К ним относятся последовательности, кодирующие гистоны, рибосомальные белки, р-РНК и т-РНК, IS-элементы, вставочные последовательности.
3) мультигенные семейства – это группы близких по структуре и функциям генов, которые «включаются» на разных этапах онтогенеза. Например b-цепь гемоглобина кодируется 7 генами, 2 из которых дефектные (псевдогены), остальные 5 включаются последовательно на разных этапах развития: в раннем эмбриогенезе, в плодном периоде (8-9 недель), в детском, юношеском и зрелом возрасте.
4) уникальные гены - специфические гены, которые кодируют синтез структурных и ферментных белков.
Структура генов эукариот. Гены эукариот имеют регуляторные элементы подобные прокариотам - промоторную и терминаторную зоны, между которыми располагается последовательность ДНК, непосредственно кодирующая белок. Регуляторные элементы генов очень важны, поскольку именно благодаря им гены «включаются» только тогда, когда есть необходимость в соответствующих белковых продуктах. Промоторная зона обеспечивает начало транскрипции и трансляции, а терминаторная зона – конец этих процессов.
В промоторах можна выделить следующие консервативные последовательности: ГЦ-мотив, ЦААТ, ТАТА, АГГАГ, инициирующий кодон АТГ (АУГ на РНК). Далее идет структурная часть гена, которая состоит из экзонов и интронов. За структурной частью гена следует зона терминатора, представленная терминирующим кодоном ТТА (ТАГ или ТГА) и терминатором. На рис. 3.1. представлены основные участки гена эукариот.
| ГЦ-мотив | ЦААТ | ТАТА | АГГАГ | АУГ | Структурная часть гена | ТАА, ТАГ, ТГА | Терми-натор |
| Палин-дром | Центр узнавания ДНК для РНК-по-лимеразы | Центр связыва-ния с РНК-полиме-разой | Центр связыва- ния с рибосо-мой | Ини-ции- рую-щий кодон | Уникальная последовательность нуклеотидов | Терми-нирую-щий кодон | Специ-фичес-кая по-следо-ватель-ность |
Рис. 3.2. Тонкая структура гена эукариот
Обозначения и пояснения к рис. 3.2.
Функции основных регуляторных элементов гена
· ГЦ-мотив – один из наиболее часто встречающихся регуляторных элементов гена. Представлен палиндромом ГГЦГГГ / ЦЦЦГЦЦ, встречается в генах общих функций, то есть тех, которые экспрессируются во всех клетках организма и играют важную роль в их жизнеобеспечении. Этот участок является, очевидно, оператором транскрипции. Присоединение к ГЦ-мотиву белка-регулятора SP1, увеличивает транскрипцию в 10-20 раз.
· ЦААТ – участок промотора гена, который, по всей видимости, распознается РНК- полимеразой перед началом транскрипции. Очевидно, этот участок выполняет ту же функцию, что у прокариот ТТГАЦА (блок Гилберта). ЦЦААТ встречается в тканеспецифичных генах, то есть тех, которые экспрессируются только в некоторых тканях и органах. Так, ген инсулина включается в основном только в клетках островков Лангерганса поджелудочной железы, ген альфа-фетопротеина - у взрослого человека только в клетках печени.
· блок Хогнеса - ТАТА (ТАТАААА или ТАТААТА), подобен блоку Прибнова (ТАТААТ) у прокариот, служит для присоединения РНК-полимеразы к ДНК в промоторной зоне, его положение в гене относительно нулевой точки начала транскрипции – (-30).
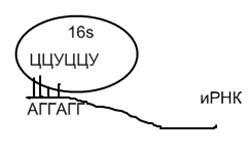
· центр связывания с рибосомой содержит редуцированную последовательность Шайна-Дальгарно АГГАГ (см. функции последовательности Шайна-Дальгарно АГГАГГ у прокариот, тема «Геномы прокариот»).
· инициирующий кодон представлен триплетом АТГ (АУГ – на РНК), транскрибируется в составе информационной РНК, с него начинается трансляция. При синтезе полипептида на рибосоме этому кодону соответствует аминокислота метионин. С метионина начинается синтез большинства белков.
· структурная часть гена – это последовательность ДНК, которая непосредственно кодирует сам белок. У эукариот, в отличие от прокариот, она не цельная, а состоит из экзонов (кодирующих участков) и интронов (вставочных некодирующих участков).
· терминирующий кодон - участок, который транскрибируется на и-РНК и обеспечивает окончание трансляции на рибосомах. На ДНК представлен нонсенс-кодонами - триплетами ТАА, ТАГ, ТГА, на РНК им соответствуют УАА, УАГ и УГА. Этим триплетам не соответствует ни одна из аминокислот, поэтому на них в рибосоме обрывается синтез полипептида.
· терминаторный участок очевидно представлен в каждом гене специфической нуклеотидной последовательностью.
В геноме эукариот обнаружили также специфические регуляторные последовательности, которые могут выступать в роли энхансеров – усилителей транскрипции, а также последовательности, которые выступают в роли сайленсеров – глушителей транскрипции. Они могут находиться на значительном удалении от гена, который регулируют, причем, одни и те же последовательности в одной клетке могут быть энхансерами, а в другой - сайленсерами. С их помощью регулируется экспрессия генов.
Обнаружены также регуляторные белки, способные связываться с промоторной зоной гена и обеспечивающие либо активацию, либо подавление транскрипции. Так, регуляторный белок SP1, связываясь с ГЦ-мотивом, может усиливать транскрипцию в 10-20 раз.
Устройство генов эукариот. Гены эукариотических организмов обладают следующими характеристиками:
- одиночные, т.е. в отличие от прокариот, не собраны в опероны;
- иногда олигомерные (представлены генами-кластерами);
- прерывистые, т.е. разделены на интроны и экзоны;
- перекрывающиеся, т.е. в пределах одного генного участка ДНК может функционировать несколько рамок считывания.
Генетический анализ у эукариот, в частности у их простейших представителей – дрожжей и нейроспоры, показал, что гены, контролирующие различные этапы одного и того же пути метаболизма, как правило, хаотично разбросаны по геному и обычно не образуют скоплений подобно оперонам бактерий. Однако было найдено несколько исключений, а именно: компактный участок ДНК у грибов контролирует 3 реакции в биосинтезе гистидина. Сходная ситуация обнаружена при изучении генетического контроля биосинтеза ароматических аминокислот (триптофана, тирозина, фенилаланина), а также – жирных кислот. У исследователей создалось впечатление, что они имеют дело с опероноподобной структурой, кодирующей мультиэнзимный комплекс. В действительности же оказалось (при использовании мутационного анализа), что у грибов все 5 этапов биосинтеза ароматических аминокислот контролирует 1 ген, продуктом которого является длинная полипептидная цепь массой 150 000 Д. Это не оперон, а ген-кластер (cluster-gene). Такие гены-кластеры довольно часто встречаются у эукариот. В качестве примеров можно привести следующие гены-кластеры:
· his 4 – ген-кластер для биосинтеза гистидина у дрожжей-сахаромицетов, кодирует единый полипептид с тремя ферментативными активностями;
· arom 1 – ген-кластер для биосинтеза ароматических аминокислот у нейроспоры, кодирует единый полипептид с пятью ферментативными активностями;
· fas 1 – первый ген-кластер для биосинтеза жирных кислот у дрожжей-сахаромицетов, кодирует полипептид с тремя ферментативными активностями
· fas 2 – второй ген-кластер для биосинтеза жирных кислот у дрожжей-сахаромицетов, кодирует единый полипептид с пятью ферментативными активностями.
Существование генов-кластеров является примером молекулярной олигомеризации. Очевидно, считывание с гена-кластера информации сразу о нескольких ферментах метаболического пути является для клетки “экономически” более выгодным, как и в оперонах прокариот. В отличие от оперона бактерий, в генах-кластерах в результате транскрипци и последующей трансляции на рибосомах синтезируется одна длинная молекула полипептида, в которой отдельные домены после пространственной укладки в третичную структуру начинают выполнять функции отдельных ферментов. В оперонах прокариот отдельные гены оперона обычно транслируются в самостоятельные белковые продукты.
Большинство же генов эукариот – одиночные, т. е в ходе эволюции эукариот происходила автономизация генов. По-видимому, это создает благоприятные условия для раздельной, а значит, и более тонкой регуляции функций отдельных генов. Напомним, что у прокариот регуляции зачастую подвержены сразу все гены оперона, за исключением аутогенного котроля, когда ген-регулятор находится среди структурных генов внутри оперона и позволяет регулировать оперон отдельными блоками.
Гены эукариот прерывистые, а именно, состоят из кодирующих участков – экзонов, и не кодирующих – интронов. Такую структуру генов называют интрон-экзонной или мозаичной структурой. Длина экзонов достигает 1000 пар нуклеотидов, а интронов – обычно 5000-20000 пар нуклеотидов. Структурная часть гена может включать 2-3 (иногда более) экзонов, разделенных длинными интронами. И хотя интронов обычно бывает немного, число их у разных видов и в разных генах может колебаться от 0 (в генах гистонов) до 51 (в структурном гене коллагена). Экзонов всегда больше, чем интронов, но на долю интронов приходится в 5-7 раз больше нуклеотидных пар, чем на долю экзонов, поскольку интроны длиннее. В зависимости от количества экзонов и интронов, а также от их длины зависит длина гена эукариот. У разных организмов она может сильно варьировать. Так, у дрозофилы средняя длина гена составляет 2 тис. п. н., а длина гена фиброина шелка у шелковичного червя достигает 16 тис. п.н.
Существование интронов в структурной части гена создает определенные трудности для реализации генетической информации, так как в транскрибируемой и-РНК оказываются «лишние» участки ДНК, которые впоследствии не должны транслироваться на рибосомах. Как же в клетке эукариот решается эта проблема? Решение было найдено американским ученым Филиппом Шарпом из Массачусетского технологического института, который открыл явление сплайсинга (от англ. to splace – сшивать без узлов).
Механизм сплайсинга. Сначала в ядре с участка хромосомы (гена) транскрибируется полностью последовательность ДНК с формированием про-и-РНК – незрелой, более длинной РНК, которая содержит как экзоны, так и интроны. Далее, когда про-и-РНК направляется из ядра в цитоплазму, при прохождении ядерной мембраны происходит сплайсинг -созревание про-и-РНК, в результате которого вырезаются интроны, а экзоны сшиваются между собой с помощью фермента, получившего название матураза. Для осуществления сплайсинга важную роль играют особые sРНК (длиной до 160 нуклеотидов), которые стягивают между собой концы интронов, что способствует их вырезанию и последующему сшиванию экзонов. В цитоплазму на рибосомы для трансляции поступает уже зрелая и-РНК, в которой нет интронов.
Интроны не всегда являются некодирующими участками. Так, у дрожжей в генах митохондрий обнаружены интроны, кодирующие синтез фермента матуразы, который участвует в вырезании интронов. В некоторых генах дрожжей обнаружены интроны, кодирующие цитохром В и т.д.
Сплайсинг осуществляется белковыми комплексами, получившими название сплайсосомы. В состав сплайсосом, помимо уже названных матураз и sРНК, входят еще белки, придающие про-и-РНК нужную конформацию. Кроме того, сплайсосома связана с ферментами, осуществляющими полиаденилирование 3/-конца и-РНК.
Типы сплайсинга: простой; альтернативный; транссплайсинг; аутосплайсинг.
Простой сплайсинг характерендля простых генов, последовательность экзонов которых предназначена для синтеза только одного белка. В таких генах экзоны занимают на ДНК всегда фиксированное положение и удаление интронов всегда ведется в четко обозначенных точках.
Альтернативный сплайсинг характерен для генных участков, на которых закодированы сразу несколько белков. При этом одни и те же участки выступают то экзонами, то интронами. Так на одном участке ДНК кодируется нейропептид гипофиза и гормон паращитовидной железы. В зависимости от вырезания тех или иных участков ДНК образуется и-РНК, кодирующая тот или иной белок. Альтернативный сплайсинг имеет место при синтезе иммуноглобулинов (антител) и при синтезе антигенов тканевой совместимости (МНС).
Транссплайсинг п роисходит, если в одну молекулу и-РНК объединяются экзоны из разных генов. Характерен для синтеза компонентов цитоскелета клетки.
Аутосплайсинг обнаружен впервые в макронуклеусе инфузорий, а позже у бактерий, дрозофил и других эукариот. Аутосплайсинг – самонарезание про-и-РНК без участия матураз и других ферментов. РНК, которая сама вырезает из себя интроны, получила название рибозим. Аутосплайсинг свидетельствует о том, что первой молекулой, несущей генетическую информацию, в эволюции была РНК. Она выполняла и генетическую и каталитическую функции, переданные позднее ДНК и белкам соответственно.
Как же в структуре генов образовались некодирующие интроны? Существует гипотеза, что еще на заре эволюции эукариот, они заражались вирусами и за счет интеграции в геном вирусной ДНК в хромосомах появилась избыточная сателлитная (эгоистическая) ДНК. Она присутствует не только в интронных последовательностях генов, но и разбросана по всей длине хромосом в виде огромных вставок некодирующих последовательностей.
У эукариот, так же как и у вирусов, встречаются перекрывающиеся гены, а именно на одном и том же участке ДНК с разных точек (и/или на разных цепях) может начинаться транскрипция с образованием разных и-РНК, кодирующих разные полипептиды.
Репликация у эукариот множественная, в каждой хромосоме существует 20-100 сайтов начала репликации и соответствующее число репликонов. Репликация в них может идти не одномоментно, однако деление клетки не начинается, пока не реплицированы все хромосомы на всем их протяжении. Подробно репликация рассмотрена в отдельной лекции (см. выше).
Транскрипция и трансляция у эукариот разобщены из-за наличия ядерной мембраны, а именно, транскрипция осуществляется в ядре, а образующаяся при этом информационная РНК должна транспортироваться из ядра в цитоплазму для последующего синтеза белка (трансляции) на рибосомах. Уже говорилось о том, что при преодолении ядерной мембраны происходит сплайсинг, т.е. созревание и-РНК. На все эти процессы необходимо время, поэтому от момента инициации транскрипции до появления белкового продукта в процессе трансляции проходит 6-24 часа. Для сравнения: у прокариот это время составляет 2-3 минуты.
|
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 1541; Нарушение авторских прав?; Мы поможем в написании вашей работы!