КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Анализ зависимости ОБЭ от ЛПЭ излучения
1. ЭФФЕКТИВНОЕ ПОПЕРЕЧНОЕ СЕЧЕНИЕ ДЕЙСТВИЯ
В некоторых случаях применимость основных идей теории мишени может быть установлена прямым наблюдением. Так, облучение делящихся клеток млекопитающих очень узким пучком протонов (диаметром около 2.5·10-4 см) с энергией 1 МэВ вызывает грубые хромосомные аберрации при пересечении хромосомы лишь несколькими десятками протонов; при прохождении через цитоплазму даже тысяч протонов никаких аберраций не возникает.
По аналогии с понятием поперечного сечения взаимодействия нейтронов можно ввести подобное понятие для ионизирующего излучения. В случае неионизирующего излучения это понятие было введено в связи с тем, что часть радиоволн огибала объект и с ним не взаимодействовала. Почему возникла необходимость ввести такое понятие для ионизирующего излучения? Если мишень действительно существует, то её поперечное сечение должно проявляться в различных опытах.
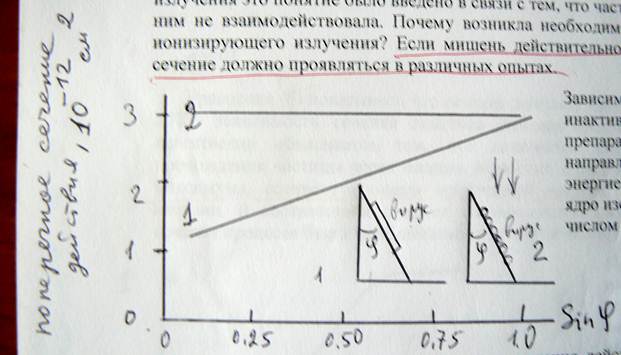
Зависимость поперечного сечения для инактивации ориентированных препаратов вируса табачной мозаики от направления пучка дейтронов с энергией 4 МэВ. Дейтрон – стабильное ядро изотопа водорода с массовым числом 2. Обозначается 2H или D или d
Рассуждения, касающиеся поперечного сечения действия, относятся прежде всего к корпускулярным излучениям. В таком случае попадание в мишень V можно связать с прохождением потока заряженных частиц через формально определяемое поперечное сечение. Обозначим это сечение действия σ. Пусть поток частиц n после облучения в дозе D. Поэтому одноударную кривую можно описать двумя различными способами:
N/No = e-VD, где v = 1 / Do, (1)
где vD – объем мишени, умноженный на дозу. Фактически это равно числу попаданий, т.е. числу частиц, пересекающих мишень. Это число может быть записано как произведение потока частиц n на эффективное сечение мишени σ, т.е.
VD = nσ (2)
Тогда S = N/No = e-VD = e-nσ
Пусть S = e-1 = e-VD= e-nσ
Отсюда
V = 1 / Do и σ = 1 / no (3)
Здесь – поток частиц, нгеобходимый для формирования одного повреждения. Из последнего выражения видно, что чем меньше поток частиц, нужный для формирования одного повреждения, тем больше сечение действия σ. Дозу можно представить через поток частиц и их ЛПЭ (L)
D = nL / ρ (4)
Действительно D = nL / ρ = [1/см2] [кэВ/мкм] [см3/г] = кэВ/г, т.е. по размерности это доза. Учитывая выражения (2) и (3), имеем
VD = nσ = VnL / ρ (5)
Отсюда
V = 1 / Do = σρ / L (6)
или для ρ = 1
σ = VL (7)
Уравнение (6) показывает, что сечение действия зависит от качества излучения, т.е. от ЛПЭ. Зависимость сечения действия (сечения инактивации в том числе) от ЛПЭ качественно объясняется тем, что количество энергии, передаваемое во время прохождения частицы через мишень возрастает с ростом ЛПЭ излучения до некоторого максимума, соответствующего критической энергии, которая должна выделиться в мишени. В соответствии с этими идеями следует ожидать, что с увеличением ЛПЭ сечение процесса будет приближаться к геометрическому сечению мишени.
Правильность этих идей в целом, а именно то, что сечение действия ведёт себя как геометрическое поперечное сечение, прекрасно иллюстрируется уже приводившимися опытами с облучением дейтронами вируса табачной мозаики.
На следующем рисунке изображена зависимость сечения инактивации гаплоидных дрожжей (верхняя часть рисунка) и их радиочувствительрности (нижняя часть рисунка) от ЛПЭ излучения.
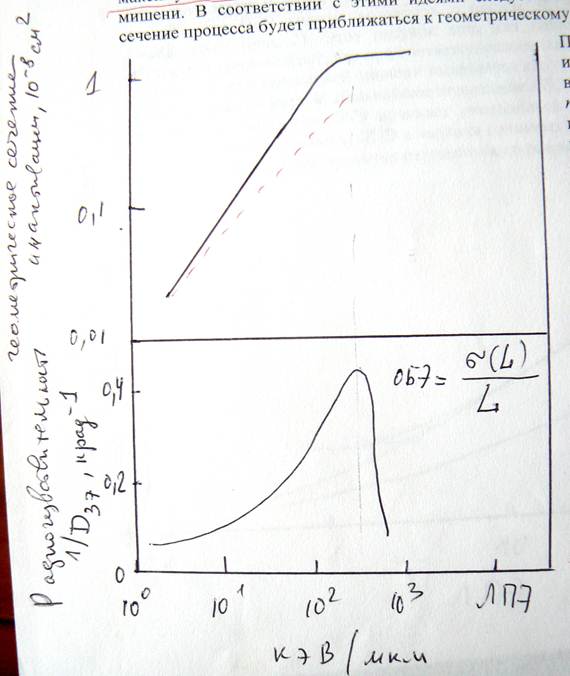
Рисунок. Зависимость сечения инактивации гаплоидных дрожжей (вверху) и их радиочувствительности (внизу) от ЛПЭ излучения.
Из этого рисунка видно, что сечение инактивации увеличивается с возрастанием ЛПЭ, причем в степени, превосходящей единицу, затем оно приближается к постоянному значению, соответствующему сечению клеточного ядра.
В общем случае для оценки биологической эффективности излучений используют понятие относительной биологической эффективности (ОБЭ), которое для исследуемого вида излучения равно отношению дозы стандартного излучения (ЛПЭ = 0,2 кэВ/мкм) к дозе исследуемого излучения при равных значениях выживаемости. Если формы кривых доза–эффект не зависят от качества излучения, тогда ОБЭ не будет зависеть от уровня биологического эффекта (выживаемости), для которого он рассчитывается.
| 
| |
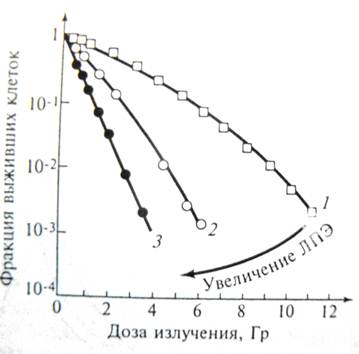
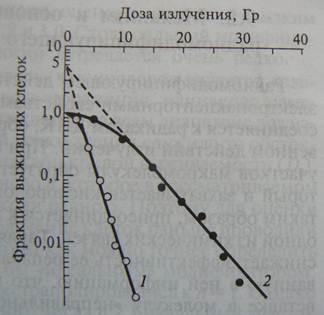
| Итак, для дрожжевых клеток форма кривой зависимости выживаемости от дозы не зависит от ЛПЭ излучения, в этом случае ОБЭ = 3,5 и не зависит от уровня важиваемости, для которого он рассчитывается ОБЭ. Другая ситуация наблюдается для культивируемых клеток млекопитающих. Сигмоидная форма кривой доза-эффект (кривая 1) после действия редкоионизирующего излучения постепенно с ростом ЛПЭ превращается в экспоненциальную. Благодаря этому, ОБЭ зависит от уровня выживаемости, для которого она рассчитывается (рис. ниже). |
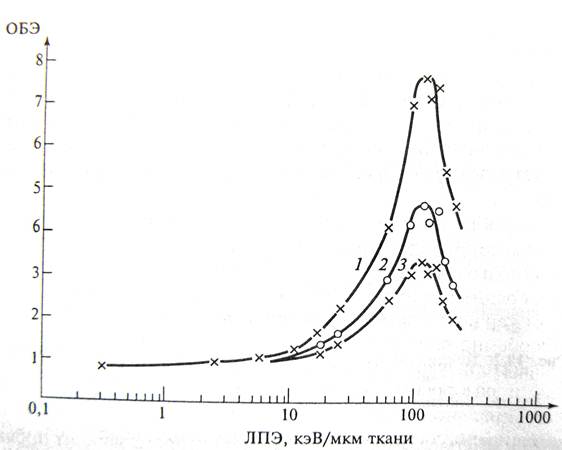
Зависимость ОБЭ от ЛПЭ и уровня выживаемости, для которого она рассчитывается для выживаемости клеток почек человека. Криая 1 – 80%, 2 – 10%, 3 – 1%
Для этого простейшего варианта – независимости формы кривой выживаемости от качества излучения, можно записать
ОБЭ(L) = Do(γ) / Do(L) = 1 / Do(L) (7)
1 / Do – радиочувствительность. Если её принять за единицу для редко ионизирующего излучения, то зависимость 1 / Do от ЛПЭ и будет зависимость ОБЭ от ЛПЭ.
Учитывая уравнение (5), получаем
V = 1 / Do = σρ / L = ОБЭ (8)
ОБЭ = σ(L)ρ / L (9)
Поскольку σ(L) возрастает до некоторого максимума (верхняя часть рис. 1) и так как σ(L) возрастает быстрее, чем L, тогда ОБЭ = max, когда σ(L) = max. Уже отсюда следует, что ОБЭ должно возрастать с ростом ЛПЭ, достигать максимума и затем уменьшаться (рис. 2). Этот рисунок дать для гаплоидных и диплоидных дрожжей. Максимум предполагает, что при соответствующем значении ЛПЭ (в данном случае ~ 130 кэВ/мкм) в чувствительной мишени выделяется как раз столько энергии, сколько нужно для инактивации клеток. Дальнейшее повышение ЛПЭ приводит к излишнему выделению энергии, за счет этого ОБЭ начинает уменьшаться. Уменьшение ОБЭ с ростом ЛПЭ означает, что при больших ЛПЭ в мишени (мишенях) образуется больше ионизаций, чем это требуется для формирования радиационного повреждения.
Чуть подробнее о причине спада ОБЭ при больших значениях ЛПЭ. Причина этого явления заключается в том, что гибель клетки происходит после поглощения достаточного количества энергии в некотором критическом объеме. Естественно, что с ростом ЛПЭ такая вероятность увеличивается. Но после некоторых величин ЛПЭ наступает насыщение, и каждая последующая частица теряет энергию уже в убитой клетке; следовательно, эффективность излучений с такой ЛПЭ падает, так кк энергия расходуется вхолостую.
Это явление наглядно представлено на следующем рисунке, где показано, что после оптимального значения ЛПЭ, когда наблюдается максимум пораженных единиц на единицу дозы (т.е. разменивается столько энергии, сколько нужно для поражения всех мишеней), наступает эффект избыточного поражения («перепоражения» – overkill).
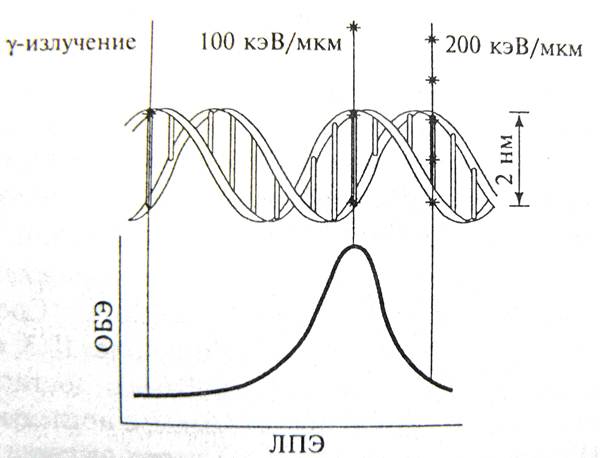
Максимальная ОБЭ наблюдается при ЛПЗ, равнолй приблизительно100 кэВ/мкм, когда достаточно двух попаданий в мишень (около 2 нм) для возникновения двойного разрыва, следствием которого является гибель клетки, мутация или онкогенная трансформация. Дальнейший рост ЛПЭ сопровождается «перепоражением».
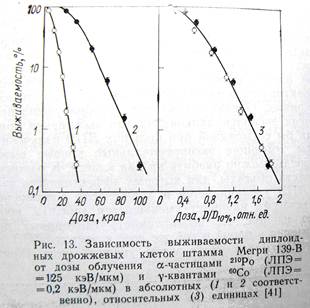
В последние годы (20–30лет назад) в нашей лаборатории были получены данные, демонстрирующие, что ОБЭ плотноионизирующих излучений определяется не только качеством излучения, т.е. ЛПЭ, но и зависят от способности биологических объектов восстанавливаться от радиационных повреждений. На большой совокупности экспериментальных данных было показано, например, что для диплоидных дрожжевых клеток, способных к пострадиационному восстановлению, ОБЭ альфа частиц (ЛПЭ – 130 кэВ/мкм) оставляет около 4–5, в то время как для диплоидных радиочувствительных мутантов ОБЭ = 1,5–2,0. Гаплоидные штаммы не обладают способностью восстанавливаться от двунитевых разрывов ДНК, поэтому они более радиочувствительны и для них ОБЭ значительно меньше, чем для диплоидных штаммов.
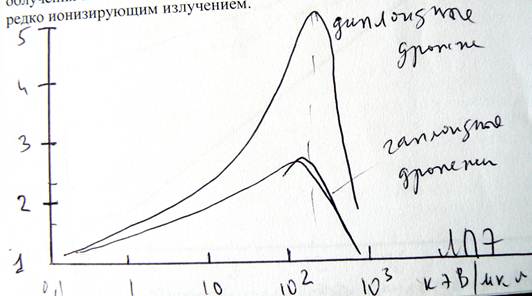
Если σ(L) = max для редко ионизирующего излучения, то ОБЭ уменьшается с увеличением ЛПЭ (см. рс. Чуть ниже)).
Итак, зависимость сечения инактивации и ОБЭ от ЛПЭ качественно объясняется тем, что количество энергии, передаваемое во время прохождения частицы через мишень возрастает с ростом ЛПЭ излучения до некоторого максимума, соответствующего критической энергии, которая должна выделиться в мишени для формирования радиационного повреждения.
2. ПРОПУСТИТЬ ТИПИЧНЫЕ ЗАВИСИМОСТИ ОБЭ ОТ ЛПЭ
Кривые выживаемости S(D) бывают экспоненциальными и сигмоидными в зави-симости характера зависимости S(D). Ниже приводятся примеры этих и более сложных кривых доза – эффект.
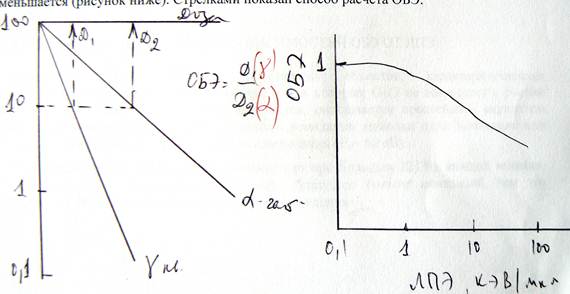
Оъекты, характеризуемые экспоненциальной зависимостью S(D) по зависимости ОБЭ от ЛПЭ подразделяются на два класса (i) объекты (бактерии, вирусы, ферменты) для которых ОБЭ не увеличивается, а только убывает с увеличением ЛПЭ, и (ii) объекты, для которых ОБЭ возрастает с увеличением ЛПЭ, достигает максимума и затем резко уменьшается (рисунок ниже). Стрелками показан способ расчета ОБЭ.
Задачи. (А) Даны Do(γ) и Do(L). Нарисовать кривые доза–эффект и рассчитать ОБЭ. (Б) По заданным двум кривым (стандартное и исследуемое излучение) рассчитать ОБЭ. (В) Дана ОБЭ и кривая доза-эффект стандартного излучения, нарисовать кривую доза–эффект для исследуемого излучения. (Г) Дана ОБЭ и кривая доза–эффект исследуемого излучения, нарисовать кривую доза–эффект стандартного излучения. (Д) Дана Do(γ) и ОБЭ. Нарисовать кривые доза–эффект. (Е) Дана Do(L) и ОБЭ. Нарисовать кривые доза–эффект.
Зависимость ОБЭ от ЛПЭ для объектов, характеризующихся сигмоидными кривыми выживаемости, аналогичным образом проходит через максимум. Такие объекты также подразделяются на два класса (i) объекты, для которых форма кривой доза – эффект не зависит от ЛПЭ и (ii) объекты, для которых сигмоидная кривая S(D) с увеличением ЛПЭ превращается в экспоненциальную. В первом случае ОБЭ плотно ионизирующих частиц не зависит от дозы, во втором – возрастает с уменьшением дозы. Отметить, что в этом случае повышение ОБЭ связано не с реальным повышением эффективности действия плотно ионизирующих излучений, а с понижением эффективности действия редко ионизирующих излучений. Дать примеры.
Задача. Рассчитать зависимость ОБЭ от дозы для экспоненциальных кривых доза – эффект (бактерии) и для кривых доза – эффект культивируемых клеток млекопитающих – сигмоидных после действия редко ионизирующего излучения и экспоненциальных после излучений с высокими ЛПЭ.
3. ПРОПУСТИТЬ КАЧЕСТВЕННОЕ ОБЪЯСНЕНИЕ ЗАВИСИМОСТЕЙ ОБЭ ОТ ЛПЭ
Зависимость ОБЭ – ЛПЭ для простейших объектов, характеризующихся экспоненциальными кривыми доза – эффект и для которых ОБЭ не возрастает с ростом ЛПЭ, а остается постоянной, а затем снижается, описывается простейшим вариантом теории попадания и мишени, когда событием попадания является одна ионизация или небольшая группа локально расположенных ионизаций (Ео~ 60 эВ).
Уменьшение ОБЭ с ростом ЛПЭ означает, что при больших ЛПЭ в каждой мишени, которую пересекает ионизирующая частица, образуется больше ионизаций, чем это требуется для формирования радиационного повреждения.
Если для наступления эффекта достаточно одной ионизации в пределах одной мишени, то (i) кривая доза – эффект должна быть экспоненциальной; (ii) эффект не должен зависеть от мощности дозы; (iii) ОБЭ не должна возрастать с ростом ЛПЭ излучения.
Результаты опытов с макромолекулами, вирусами и бактериофагами удовлет-ворительно описываются одноударной моделью, в которой для поражения мишени достаточно одной пары ионов, образованных в пределах этой мишени.
Задача. Дать пять кривых выживаемости бактерий для 60Со (ЛПЭ = 0,2 кэВ/мкм), нейтронов (ЛПЭ =!0 и 50 кэВ/мкм), α-частиц (ЛПЭ = 130 кэВ/мкм) и ядер углерода (ЛПЭ = 400 кэВ/ мкм). Нарисовать зависимость ОБЭ от ЛПЭ.
Увеличение эффективности действия излучения (ОБЭ) с ростом ЛПЭ излучения для экспоненциальных кривых допускает различные интерпретации, общим требованием для которых является образование одной ионизирующей частицей нескольких пар ионов в одной или нескольких мишенях или одной пары ионов в нескольких мишенях. Главная проблема таких моделей – умение сосчитать вероятность Pk образования k пар ионов в гипотетических мишенях.
Многоударная модель (много попаданий в одну мишень) качественно объясняет зависимость формы кривой выживаемости от ЛПЭ излучения. В случае редко ионизирующего излучения необходимо много треков частиц для формирования многих попаданий (кривая доза – эффект сигмоидна), в то время как одна плотно ионизирующая частица формирует сразу много попаданий, и кривая доза – эффект становится экспоненциальной.
Для объектов, у которых сигмоидная кривая доза – эффект не зависит от ЛПЭ излучения, каждая частица, попадающая в эффективный объем, должна пересекать ограниченное число чувствительных участков. В этом случае с ростом ЛПЭ будет увеличиваться вероятность повреждения одного чувствительного участка (увеличение ОБЭ). Для инактивации клетки необходимо несколько попаданий ионизирующих частиц даже при очень высоких ЛПЭ, вследствие чего кривые выживаемости сохраняют сигмоидную форму при увеличении ЛПЭ частиц.
4. ПРОПУСТИТЬ ИНТЕРПРЕТАЦИЯ ОПЫТОВ С ДРОЖЖЕВЫМИ КЛЕТКАМИ
Кривые выживаемости для гаплоидных дрожжевых клеток имеют экспоненциальную форму независимо от качества излучения. Поэтому ОБЭ не зависит от уровня выживаемости, для которого она рассчитывается. Кривые выживаемости для диплоидных дрожжевых клеток имеют сигмоидную форму, причем такая форма сохраняется, в отличие от клеток млекопитающих, и при высоких значениях ЛПЭ, хотя эффективность облучения плотно ионизирующими излучениями в 4–5 раз более высокая, чем облучение редко ионизирующим излучением.
На рисунке (предыдущая страница) приведены зависимости ОБЭ от ЛПЭ одновременно для гаплоидных (нижняя кривая) и диплоидных (верхняя кривая) дрожжевых клеток. Видно, что качественно изменение этих кривых происходит одинаково с ростом ЛПЭ излучения. Сначала ОБЭ возрастает, но, начиная с ЛПЭ ~ 120–130 кэВ/мкм, ОБЭ уменьшается. Главное различие кривых зависимости ОБЭ от ЛПЭ для гаплоидных и диплоидных клеток заключается в величине максимума. Видно, что абсолютное значение ОБЭ существенно выше для диплоидных (ОБЭ = 5), чем гаплоидных дрожжей (ОБЭ = 2).
Шуточная задача. Во сколько раз ОБЭ для диплоидных дроржжей больше, чем для гаплоидных? Сравнение – во сколько раз на 4-й этаж подниматься выше, чем на второй?
Главная причина, объясняющая большую эффективность плотно ионизирующих излучений с позиций теории попадания и мишени заключается в большей величине энергии, необходимой для формирования первичного повреждения. Причина такого различия может заключаться в большей способности диплоидных дрожжей к пострадиационному восстановлению. Благодаря наличию двойного набора хромосом, они могут восстанавливаться как от одиночных, так и от двойных разрывов ДНК, в то время как гаплоидные дрожжи могут репарировать только одиночные разрывы ДНК.
Следовательно, мы приходим к выводу, что ОБЭ плотно ионизирующих излучений определяется не только физическими характеристиками излучения, но и биологическими свойствами объекта – его способностью клеток восстанавливаться от радиационных повреждений. Большой комплекс экспериментальных данных с радиочувствительными мутантами дрожжей подтверждает эту точку зрения. Главный результат этих экспериментов заключался в том, что даже для диплоидных дрожжевых клеток, утративших свою способность к восстановлению, ОБЭ существенно меньше, чем для штамма «дикого» типа, сохранившего такую способность.
Остается разобрать вопрос, почему у диплоидных дрожжевых клеток форма кривой выживаемости не зависит от ЛПЭ излучения, в то время для культивируемых клеток млекопитающих (тоже диплоидных) сигмоидная форма кривой выживаемости после действия редко ионизирующего излучения трансформируется в экспоненциальную после действия излучений с высокими ЛПЭ?
Отметим, что дрожжевые клетки, как правило, в радиобиологических экспериментах облучают в стационарной стадии роста, а клетки млекопитающих – в логарифмической стадии роста. Почему это происходит так, поясним чуть позже. Ниже на рисунке приводится динамика роста дрожжевых клеток в питательной среде.
Видно, что увеличение концентрации клеток происходит экспоненциально – в логарифмическом масштабе рост клеток характеризуется прямой линией. Это и есть логарифмическая стадия роста. Видно также, что примерно после 30 часового культивирования в питательной среде концентрация клеток выходят на плато и дальше остается постоянной. Это и есть стационарная фаза роста. Максимально достигаемая концентрация клеток (М-концентрация) не зависит от начальной концентрации клеток. Дрожжевые летки в стационарной стадии роста имеют 100% эффективность роста, очень гомогенны по своим размерам, чувствительности к действию ионизирующего излучения и других повреждающих факторов. Более сложная ситуация наблюдается для дрожжевых клеток, облученных в логарифмической стадии роста. Из рисунка на странице видно, что кривые доза–эффект клеток, облученные в логарифмической стадии роста, имеют резистентный «хвост», связанный с гетерогенностью облучаемой популяции. Эта гетерогенность объясняется тем, что часть гаплоидных клеток имеют двойной набор хромосом, а диплоидных – тетраплоидных. В этой части популяции хромосомы удвоились, но не успели перейти в дочернюю почку. Таким образом, при облучении популяции гаплоидных дрожжей в логарифмической стадии, часть этих клеток являются фактически диплоидными, имеющие двойной набор хромосом и характеризующиеся повышенной устойчивостью к действию ионизирующего излучения. Именно этой повышенной резистентностью диплоидных клеток и объясняется наличие резистентного хвоста на кривой выживаемости. Именно по этой причине дрожжевые клетки в радиобиологических экспериментах не облучают в логарифмической стадии роста.
В таком случае возникает вопрос, почему культивируемые клетки млекопитающих облучают в логарифмической, а не в стационарной стадии роста. Это обусловлено совершено другой причиной. Дело в том, что клетки млекопитающих в стационарной стадии роста характеризуются пониженной эффективностью роста, которая может достигать в некоторых случаях 30–70%. Следовательно, клетки млекопитающих, достигших стационарной стадии роста, уже несут в себе какие-то повреждения, и их кривые выживаемости часто имеют экспоненциальный характер. Именно по этой причине в радиобиологических экспериментах клетки млекопитающих облучают в стационарной стадии роста.
Известно, что с ростом ЛПЭ излучения эффективное поперечное сечение инактивации возрастает, стремясь к постоянному значению, причем в случае прокариотов это значение близко к геометрическому сечению самого объекта, а в случае эукариотов – к сечению ядра клетки. Это означает, что гипотетические чувствительные участки (мишени) распределены в первом случае по всему объекту, а во втором – по объему ядра.
Тогда превращение сигмоидной кривой выживаемости в экспоненциальную с ростом ЛПЭ излучения можно объяснить равномерным распределением чувствительных участков (участки хромосом) по всему объему мишени (клеточное ядро). В этом случае при действии редко ионизирующего излучения необходимо прохождение нескольких ионизирующих частиц для формирования критического числа повреждений, и поэтому кривая выживаемости будет сигмоидной. Каждое прохождение плотно ионизирующей частицы будет сопровождаться поражением достаточного количества чувствительных участков, и поэтому кривая выживаемости будет экспоненциальной, что и наблюдается для клеток млекопитающих. Для диплоидных дрожжевых клеток, облученных в логарифмической стадии роста, также наблюдается подобное превращение сигмоидной формы в экспоненцильную, но если принимать во внимание только начальный участок кривой доза-ффект и не принимать во внимание резистентный хвост, природу возникновения которого мы только что обсудили.
При облучении дрожжевых клеток в стационарной стадии роста кривая выживаемости остается сигмоидной даже при очень высоких значениях ЛПЭ. В этом случае форма мишени должна быть такой, чтобы даже плотно ионизирующая частица пересекала бы ограниченное число участков внутри ядра, т.е. чувствительные участки должны быть распределены не равномерно по всему ядру, а сконцентрированы в каких-то местах. В качестве рабочей гипотезы можно полагать, что у диплоидной дрожжевой клетки мишень имеет форму сферического слоя, диаметр которого равен диаметру клеточного ядра, а толщина составляет несколько десятков ангстрем.
Таким образом, анализ с позиций принципа попаданий кривых выживаемости диплоидных дрожжей и клеток млекопитающих показал, что для интерпретации результатов, получаемых с этими объектами, приемлема многоударная модель, в которой событием попадания (ударом) является локальное образование одной ионизирующей частицей нескольких ионов в одной из чувствительных структур, предположительно хромосоме. В зависимости от стадии клеточного роста чувствительные структуры могут располагаться либо на периферии клеточного ядра, образуя тонкий сферический слой, либо равномерно занимать внутренние области клеточного ядра.
|
Дата добавления: 2014-01-05; Просмотров: 2590; Нарушение авторских прав?; Мы поможем в написании вашей работы!