КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Транспорт липидов
|
|
|
|
Ресинтез жирных кислот в стенке кишечника.
Из всосавшихся продуктов гидролиза в стенке кишечника происходит ресинтез специфичных для данного организма. Триглицеридов и фосфоглицеридов. При этом происходит последовательное присоединение к моноглицеридам активированных жирных кислот.
При синтезе фосфолипидов глицерин фосфорилируется фосфатглицеролкиназой, превращается в глицерол-3-фосфат, который затем взаимодействует с активированными жирными кислотами, образуя фосфатидную кислоту.
Ресинтезированные триацилглицерины, фосфолипиды, холестерин и его эфиры в эпителиальных клетках кишечника соединяются |с небольшим количеством белка и образуют хиломикроны (ХМ- частицы диаметром d=0,1-5 мкм). ХМ поступают в лимфотическую систему, а оттуда – через грудной проток в кровь. Из-за большого размера хиломикроны не могут сразу всосаться в кровеносные капилляры. В хиломикронах белки и полярные части фосфолипидов расположены снаружи, а триглицериды и холестерин внутри.
По крови хиломикроны транспортируются к печени и жировой ткани. На поверхности клеток под действием фермента липопротеиназа происходит их гидролиз с образованием глицерина и жирных кислот. Часть жирных кислот поступает в клетки жировой ткани и там откладываются в виде триглицеридов, а часть образует комплексы с альбуминами сыворотки крови и оком крови разносится к тканям. В клетках тканей комплекс распадается и жирные кислоты подвергаются биологическому окислению, обеспечивая клетку энергией, либо используется для синтеза триглицеридов жировой ткани, а также липопротеинов, фосфолипидов, стеридов и др. соединений.
Жирные кислоты, как и глюкоза, является основным энергетическим материалом в организме. При повышенных энергозатратах происходит мобилизация жирных кислот из жировых депо. При этом триглицериды резервного жира последовательно расщепляются под действием фермента триглицеридлипаза (активизируется адреналином и др. гормонами), ди- и моноглицеридлипазами с образованием глицерина и жирных кислот, которые в виде комплексов с альбуминами кровью переносятся к тканям где подвергаются процессам распада или биосинтеза.
|
|
|
Тема: ОРГАНИЧЕСКИЕ КИСЛОТЫ.
Органические кислоты широко распространены в растительном мире. Они содержатся во всех растениях и часто накапливаются в больших количествах в семенах, стеблях, листьях, корнях или плодах. Органические кислоты образуются в результате многих процессов обмена веществ в растениях, среди которых основным следует считать дыхание.
Органические кислоты участвуют в построении молекул ряда сложных веществ — жиров, производных сахаров, витаминов и других биологически активных соединений. В растениях кислоты находятся в свободном состоянии, а также в виде кислых и нейтральных солей. Во многих плодах и ягодах большая часть приходится на свободные кислоты и лишь незначительное количество представлено солями. В некоторых растениях (например, щавеле, бегонии, суккулентах) много свободных кислот содержится и в листьях. В связанном состоянии органические кислоты обнаружены в больших количествах в листьях бобовых растений, где на их долю может приходиться до 25% сухого вещества.
При определениях содержания органических кислот или их солей следует различать: 1) общую кислотность, или общее содержание кислоты, понимая под этим общее количество анионов и недиссоциированных молекул кислоты; 2) концентрацию водородных ионов, часто обозначаемую «истинная кислотность»; 3) титруемую кислотность; под этим часто понимают концентрацию «свободной» кислоты. Однако это справедливо только для одноосновных кислот. У двуосновных кислот часть общей кислотности, которую можно титровать щелочью, состоит из двух фракций: недиссоциированной кислоты и одновалентных кислотных ионов.
|
|
|
При анализах органические кислоты можно экстрагировать из свежих, замороженных или высушенных растительных тканей. Однако следует иметь в виду, что при высушивании в условиях повышенной температуры могут происходить изменения в содержании органических кислот (потери летучих кислот или их эфиров, взаимные превращения кислот, взаимодействие их с углеводами и т. д.), поэтому для определения органических кислот сушить материал лучше при комнатной температуре в вакууме или применять высушивание лиофилизацией.
Для экстракции кислот можно использовать воду или органические растворители, из которых чаще всего — эфир. Однако эфир растворяет свободные кислоты, но не соли органических кислот, поэтому для экстракции солей экстрагируемый материал надо предварительно подкислять минеральной кислотой.
При экстракции водой из растительного материала, кроме кислот, извлекается много сопутствующих веществ— сахаров, пектиновых веществ, аминокислот, белков, которые перед количественным определением необходимо тщательно удалять из раствора. Удаляют эти вещества или экстракцией и осаждением, или при помощи катионо- и анионообменных смол. Эфир не извлекает углеводов, аминокислот и белков, но растворяет жиры и липоиды.
Цикл трикарбоновых кислот (цикл Кребса)
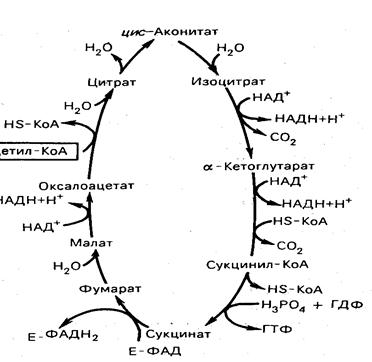
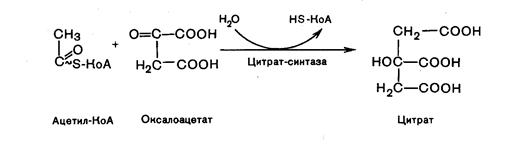
Первая реакция катализируется ферментом, цитратсинтазой, при этом ацетильная группа ацетил-КоА конденсируется с оксалатом, в результате образуется лимонная кислота:
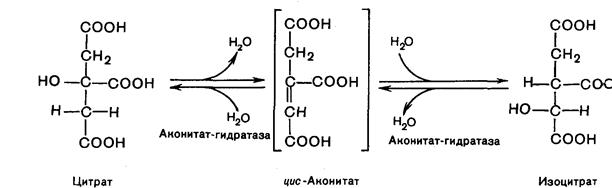
Вторая реакция. Образовавшаяся лимонная кислота подвергается дегидрированию с образованием цис-аконитовой кислота, которая присоединяя молекулу воды, переходит в изолимонную кислоту (изоцитрат):
Третья реакция лимитирует скорость цикла Кребса. Изолимонная кислота дегидируется в присутствии НАД-зависимой изоцитратдегидрогеназы:
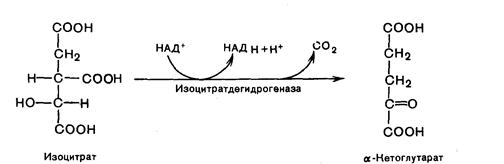
В ходе изоцитратдегидрогеназной реакции изолимонная кислота одновременно декарбоксилируется. Фермент для проявления своей активности нуждается в ионах Мg2+ или Мn2+.
|
|
|
Четвертая реакция. Происходит окислительное декарбоксилирование α-кетоглутаровой кислоты с образованием высокоэнергетического соединения сукцинил-КоА. Механизм этой реакции сходен с окислительного декарбоксилирования пирувата до ацетил-КоА. В реакции принимают участие 5 коферментов: ТПФ, амид липоевой кислоты, НS-КоА, ФАД и НАД+.
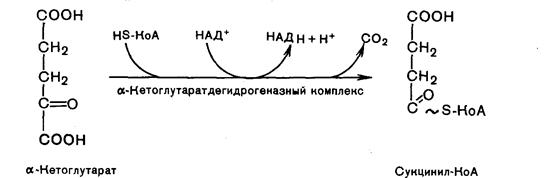
Пятая реакция катализируется ферментом сукцинил-КоА-синтетаза. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Одновременно происходит образование высокоэргической фосфатной связи ГТФ за счет высокоэргической тиоэфирной связи сукцинил-КоА:
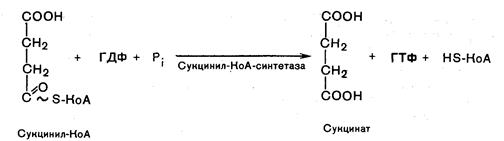
Шестая реакция. Сукцинат дегидрируется в фумаровую кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой, в молекуле которой с белком прочно (ковалентно) связан кофермент ФАД, а сам фермент прочно связан с внутренней митохондриальной мембраной:
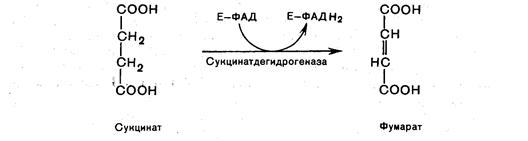
Седьмая реакция. Фумаровая кислота под действием фермента фумаратгидратазы (фумаразы) гидратируется и получается L-яблочная кислота (малат)
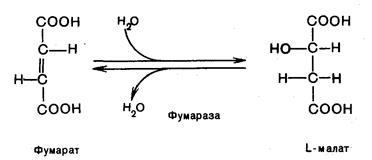
Восьмая реакция. Происходит окисление L-малата в оксалоацетат под действием митохондриальной НАД-зависимой малатдегидрогеназы:
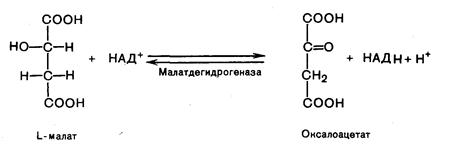
Таким образом, за один оборот цикла состоящего из 8 ферментативных реакций, происходит полное окисление одной молекулы ацетил-КоА с образованием 12 молекул АТФ.
Энергетика ЦТК:
Образуется 3 НАДН2, 1 ФАДН2 и ГТФ.
Каждая молекула НАДН2 дает в системе тканевого дыхания 3 молекулы АТФ, ФАДН2 – 2 молекулы АТФ и ГТФ – 1 молекулу АТФ.
3*3АТФ + 2АТФ + 1АТФ = 12АТФ
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 383; Нарушение авторских прав?; Мы поможем в написании вашей работы!