КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Трудностью регулирования осмотического давления у пресноводных организмов объясняется их видовая бедность по сравоению с обитателями моря
|
|
|
|
Водный обмен и осморегуляция у земноводных.
Солевой обмен у наземных позвоночных.
Адаптации у животных к переувлажнению и недостатку влаги.
Адаптации к аридности климата.
Экологические группы растений по отношению к воде (влажности среды).
Наземный тип водного обмена у животных.
Чётко выраженную приспособленность к сезонному ритму влажности проявляют и некоторые многолетние растения, называемые эфемероидами или гео эфемероидами. Они могут задержать своё развитие в случае неблагоприятных условий влажности до тех пор, пока она не станет оптимальной или, как эфемеры, пройти весь цикл его в чрезвычайно сжатые ранневесенние сроки. Это типичные растения южных степей – тюльпаны, гиацинт степной, птицемлечники и др.
Большинство водных животных пойкилотермны, и температура их тела зависит от температуры окружающей среды. У гомойотермных же млекопитающих (ластоногие, китообразные) образуют мощный слой подкожного жира, выполняющий теплоизоляционную функцию.
Для водных животных имеет значение давление среды обитания. В связи с этим выделяются стенобатные животные, не выдерживающие больших колебаний давления, и эврибатные, обитающие и при высоком и при низком давлении. Голотурии живут на глубине от 100 до 9000 м, а многие виды раков, погонофоры, морские лилии располагаются на глубинах от 3000 до 10 000 м. У таких глубоководных животных отмечаются специфические черты организации: увеличение размеров тела, исчезновение или слабое развитие известкового скелета; часто редукция органов зрения; усиление развития осязательных рецепторов; отсутствие пигментации тела или, наоборот, тёмная окраска.
|
|
|
Проследим на примере рыб, как осуществляется осморегуляция животных в морских и пресных водах. Пресноводные рыбы избыток воды удаляют усиленной работой выделительной системы, а соли поглощают через жаберные лепестки. Морские рыбы, наоборот, вынуждены пополнять запасы воды и поэтому пьют морскую воду, а излишки поступающих с ней солей выводят из организма через жаберные лепестки.
Экологические варианты осморегуляции.
Кислород и углекислый газ.как экологические факторы
Кислород и углекислый газ. Механизмы газообмена у растений и животных. Газообмен в водной среде. Принцип водного дыхания. Адаптация к изменениям содержания кислорода в воде.
Перечисленные особенности водной среды обусловливают соответствующее строение и функции дыхательной системы. У позвоночных животных, отличающихся относительно высоким уровнем обмена веществ, простой контакт дыхательной поверхности с водной средой (кожа, пассивно свешивающиеся в воду жабры), свойственный некоторым беспозвоночным, не в состоянии обеспечить необходимую скорость поступления кислорода в организм. У всех рыб жаберный аппарат устроен так, что вода активно прокачивается сквозь систему жаберных лепестков, через поверхность которых происходит газообмен; в менее эффективной форме это выражено у круглоротых. У костных рыб движения ротового и жаберного аппаратов сочетают нагнетательный (ротовая полость) и всасывающий (жаберная полость) принципы, что обеспечивает интенсивное продвижение воды сквозь жабры. У акуловых рыб благодаря отсутствию жаберной крышки этот механизм выражен слабее; при быстром движении, сопряженном с большими затратами энергии, для прокачивания воды через жабры используется само движение: рыба плывет с открытым ртом, и вода проталкивается через жабры тем интенсивнее, чем выше скорость движения. Такой же «пассивный» тип жаберной вентиляции отмечен и у быстро плавающих костистых рыб (например, у тун цов). Видимо, при большой скорости этот тип создания дыхательного потока воды оказывается более экономным. В опытах с полосатым окунем Moronesaxatilis и луфарем Pomatomussaltatrix выяснено, что при крейсерских скоростях оба вида используют как активную, так и пассивную жаберную вентиляцию, а при наиболее форсированном движении — только пассивную. Показано, что это дает значительный (до 30%) энергетический выигрыш при критических скоростях (Fredman, 1979).
|
|
|
Во всех описанных случаях вода проходит сквозь жабры водном направлении; движений типа «вдох — выдох» у водных позвоночных нет.
Соотношение дыхательных движений ротовой полости и жаберной крышки таково, что вода прокачивается через жабры практически непрерывно (рис. 11): давление в ротовой полости на протяжении почти всего дыхательного цикла выше, чем в жаберной.
Извлечение О2 из воды в жабрах костистых рыб усиливается и вследствие того, что направление движения потока воды сквозь жаберные лепестки и тока крови в капиллярах, проходящих по вторичным жаберным пластинкам, противоположны (рис. 12). Такая противоточная система способствует тому, что на протяжении всей длины жаберной пластинки сохраняется градиент концентрации О2 (и СО2) в крови и воде, благодаря чему процесс диффузии идет непрерывно, и отходящая от жабр кровь имеет почти тот же уровень насыщения О2, что и поступающая в жабры вода. Математический анализ принципов газообмена показал, что именно противоточная система обеспечивает наибольшую эффективность утилизации O2 (Piiper, Sheid, 1972).

идет непрерывно, и отходящая от жабр кровь имеет почти тот же уровень насыщения О2, что и поступающая в жабры вода. Математический анализ принципов газообмена показал, что именно про тивоточная система обеспечивает наибольшую эффективность утилизации O2 (Piiper, Sheid, 1972).
Благодаря всем рассмотренным особенностям дыхания некоторые костистые рыбы могут извлекать до 85% О2, растворенного в омывающей жабры воде. Велика у костистых рыб и степень утилизации О2, поступившего в кровь: она в 2,5—3 раза выше, чем у млекопитающих.
У хрящевых рыб эффективность извлечения О2 из воды составляет 70—77% (при более быстром протоке 40—50%), что вполне сопоставимо с эффективностью извлечения О2 у костных рыб. От метим для сравнения, что у миног с их малоэффективным типом жаберного аппарата эта величина составляет 10—25%.
Приспособления к изменению содержания кислорода в воде, Особенности строения жабр (число и величина жаберных лепестков, количество вторичных жаберных пластинок), а соответственно и общая площадь дыхательной поверхности довольно широко варьируют у разных видов рыб. Эти показатели, характеризующие уровень дыхательной функции, хорошо коррелируют с экологическими особенностями разных видов, в том числе с их способностью к быстрому движению. Так, у быстро плавающей макрели общая жаберная поверхность в пять с лишним раз больше, чем у придонной рыбы-удилыцика, почти не совершающего активных движений. Дыхательная поверхность жабр трески и щуки примерно в полтора раза больше, чем у камбалы. Подобные адаптации отмечены и в связи с кислородным режимом водоемов: у ряда видов, обитающих в условиях пониженного содержания О2, отмечается удлинение жаберных лепестков, увеличение числа жаберных пластинок и возрастание суммарной дыхательной поверхности. Например, у циркумполярного вида Scopelarchusguentheri особи из северной части Индийского океана, где концентрация О2 в подповерхностных водах понижена, отличаются более длинными жаберными лепестками и соответственно большим числом расположенных на них вторичных пластинок.
|
|
|
Адаптации к колебаниям концентрации О2 в дыхательной среде осуществляются и на физиологическом уровне. В эксперименте за регистрировано, что на снижение количества О2 рыбы реагируют компенсаторным увеличением частоты дыхательных движений и (или) увеличением объема воды, пропускаемой через жаберный аппарат. При гипероксии дыхание, напротив, замедляется. Такая реакция имеет довольно общий характер: она отмечена у разных видов хрящевых и костных рыб, равно как и у круглоротых.
Обычно реакция гипервентиляции сопровождается замедлением сердечного ритма (брадикардия), что отражает некоторое снижение уровня метаболизма, а соответственно и потребности в О2. Можно полагать, таким образом, что «борьба за кислород» в условиях его временной недостаточности идет по двум путям: повышения интенсивности работы газообменного аппарата и некоторого снижения затрат СО2 в организме.
Физиологические механизмы регуляции дыхания у рыб изучены еще недостаточно. Ритм дыхательных движений поддерживается стимулами, идущими от клеток дыхательного центра, расположенных в продолговатом мозге. На фоне автономной активности этих клеток действуют механизмы, изменяющие ритм дыхания под влиянием динамики концентрации О2 и СО2 в крови. Эти же механизмы определяют изменение дыхательных функций в связи с колебаниями содержания О2 в воде: показано, что рыбы реагируют не на состав воды, а на изменение напряжения О2 в крови. В опытах ответ на изменение напряжения О2 в воде возникает с задержкой, величина которой хорошо коррелирует с временем, не обходимым для того, чтобы измененный состав воды успел вызвать соответствующий сдвиг содержания О2 в крови. Локализация кислородных рецепторов точно не выяснена. Скорее всего, они расположены не в самих жабрах (брадикардия возникает и при отключении тока крови через жабры), а на их поверхности, в спинной аорте, в ротовой полости или же в ложножабрах (Randall, Smith, 1967; Bamford. 1974).
|
|
|
Существенные адаптации к колебаниям напряжения О2 обнаруживаются в транспортной функции крови. Способность крови к переносу О2 обеспечивается, во-первых, растворением его в плаз ме, во-вторых — обратимым связыванием О2 с дыхательным пигментом крови (у позвоночных это гемоглобин). Количество О2, которое может быть растворено в плазме, ограничено; этот путь транспорта у большинства рыб имеет подчиненное значение. Известно лишь небольшое число форм, преимущественно антарктических (сем. Chaenichthydae), кровь которых полностью лишена гемоглобина, и весь О2 транспортируется только в плазме. Исследования, проведенные на одном из представителей этого семейства — ледяной рыбе Chaenocephalusaceratus, показали, что этот вид обитает в хорошо аэрированных холодных водах, ведет довольно пассивный образ жизни и обладает низким уровнем обмена веществ. Для него характерны обильное снабжение сосудами (васкуляризация) жабр, повышенная роль кожного газообмена, расширение сердца и увеличенная скорость кровотока в жаберных сосудах. Однако даже при этих свойствах устойчивость к гипоксии у ледяной рыбы (и у других рыб, лишенных гемоглобина) существенно ниже, чем у видов, имеющих гемоглобин.
Способность крови транспортировать кислород в связанном с гемоглобином состоянии зависит от общего количества гемоглобина (и соответственно от числа эритроцитов), а также от его химических свойств, в частности от сродства гемоглобина к кислороду, определяющего способность гемоглобина к насыщению кислородом при данном его парциальном давлении.
Кривая диссоциации оксигемоглобина (рис. 13), показывающая степень насыщения гемоглобина кислородом при различном его парциальном давлении, у наземных позвоночных обычно имеет сигмоидную форму, а у рыб — чаще гиперболическую, что свидетельствует о большем сродстве к кислороду. На этой кривой в качестве стандартных параметров приняты две точки: напряжение кислорода (в миллиметрах ртутного столба), при котором гемо глобин насыщается на 100% («зарядное напряжение»), и напряжение, при котором оксигемоглобин отдает 50% кислорода («разрядное напряжение», или «напряжение полунасыщения»).
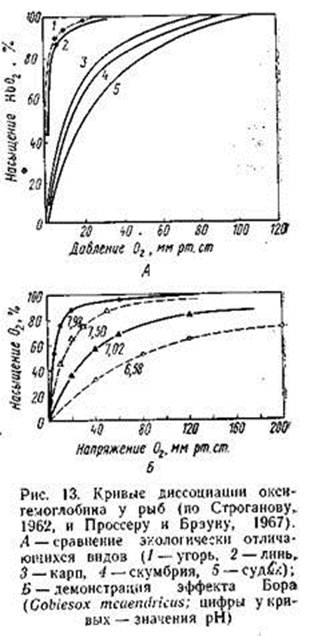
При повышении сродства гемоглобина к кислороду кривая сдвигается влево (насыщение происходит при меньшем РО2). Этот эффект может зависеть от условий, в которых происходит газообмен; в частности, связи гемоглобина с кислородом ослабевают при повышении температуры. Большое значение имеет также величина рН: при повышении кислотности сродство гемоглобина к кислороду уменьшается и кривая диссоциации сдвигается вправо («эффект Бора»). Эффект Бора имеет очень большое значение при тканевом газообмене: имен но в тканях в кровь поступает большое количество СО2, рН понижается и гемоглобин начинает легче отдавать О2 тканям.
Свойства гемоглобина у разных видов рыб могут довольно широко варьировать, отражая приспособления к кислородному режиму водоемов, в которых они обитают. В частности, у рыб, живущих в стоячих, богатых органикой водоемах, сродство гемоглобина к кислороду, как правило, выше, чем у обитателей быстротекущих вод или у неглубоководных морских рыб, которые не лимитированы количеством растворенного в воде кислорода (табл. 9).

Гематологические отличия связаны и с подвижностью: обитатели хорошо аэрированных водоемов обычно высокоподвижны; при этом пониженное сродство гемоглобина к кислороду, при котором он быстрее отдается в ткани, биологически оправдано. Рыбы из водоемов с низким содержанием кислорода, напротив, обычно ведут малоподвижный образ жизни, при котором быстрота отдачи. кислорода в ткани не столь существенна. Кроме того, у таких видов обычно хорошо выражен эффект Бора, облегчающий процесс тканевого газообмена.
Экологические корреляции дыхательных свойств крови с естественными условиями кислородного режима прослеживаются и в пределах одного вида, в частности, в зависимости от возрастных особенностей. Показано, что личинки миног Lampetrafluviatilis отличаются более высоким сродством гемоглобина к кислороду (Р50 = 0,25 кПа), чем взрослые формы (Р50=1,43 кПа), что связано с обитанием личинок в грунте, т. е. в условиях затрудненного доступа кислорода (Bird et al, 1976).
В реактивных ответах на гипоксию большую роль играет число эритроцитов: при появлении признаков кислородной недостаточности происходит выброс депонированных эритроцитов в кровь, что увеличивает ее кислородную емкость. Отмечено, что при этом рыбы, в естественных условиях испытывающие недостаток кисло рода, резче реагируют на возникновение гипоксии (табл. 10).
Газообмен у ныряющих животных.
Своеобразные условия газообмена складываются у наземных животных, вторично перешедших к водному образу жизни или эко логически связанных с регулярным нырянием либо (как многие птицы) погружением головы в воду. Все эти виды имеют легочный тип дыхания, у всех процесс внешнего газообмена происходит в нормальных атмосферных условиях. Однако во время пребывания под водой резко изменяются условия снабжения тканей кислородом: поступление его в организм прекращается и возникает состояние функциональной асфиксической гипоксии. У наиболее специализированных ныряльщиков остановка дыхания может быть до вольно длительной, хотя у подавляющего большинства видов продолжительность пребывания под водой не превышает нескольких минут.
Среди рептилий наибольшая (более часа) длительность нахождения под водой регистрируется у водных черепах и морских змей. Так, тихоокеанская зеленая черепаха Cheloniamtjdas выдерживает погружение до 5 ч, а у бородавчатых змей Acrochordusgranulatus время естественного погружения доходит почти до 2 ч. У других водных рептилий продолжительность ныряния обычно не превышает 20-25 мин. Надо отметить, что у водных рептилий значительная часть газообмена при нырянии совершается водным путем через кожу и слизистые оболочки ротовой и анальной полостей (Хозацкий, 1967; Graham, 1974).
Среди млекопитающих наиболее длительное пребывание в погруженном состоянии отмечено у вторичноводных форм - китообразных и ластоногих. Продолжительность ныряния у разных видов китов колеблется от 30 мин (финвал Balaenopteraphysalus, горбатый кит Megapteranovaeangliae) до 2 ч (бутылконос Hyperoodonampulatus). Дельфины совершают длительные серии относительно коротких ныряний; у касатки, например, продолжительность от дельного заныривания обычно не превышает 4-5 мин. Ластоногие чаше всего находятся под водой 15-30 мин (тюлень Уэддела - до 45 мин). Такие полуводные млекопитающие, как ондатра, бобр, выдерживают под водой не более 15 мин, а бегемоты, ламантины, дюгони, хотя и проводят в водоеме весьма длительное время, в погруженном состоянии могут вынести лишь несколько (обычно не более 10) минут.
Птицы по длительности погружения в большинстве случаев уступают водным млекопитающим. Длительность естественного погружения у них чаще всего меньше минуты. Ныряние буревестников, некоторых утиных, оляпки длится примерно до 15 с, у других утиных, а также у поганок - до 30 с. При насильственном погружении в воду (хотя бы только головы) тупик способен задержать дыхание на 4 мин, а чистики и кайры - даже до 12 мин. Пингвины (Pygoscelispapua, P. adeliae, Eudipteschrisolophus) при есте ственном нырянии находятся под водой 5-7 мин, а императорский пингвин Aptenodytesforsteri - до 18 мин.
Ныряние вызывает довольно быстрое уменьшение содержания О2 и нарастание концентрации СО2 в крови. Устойчивость ныряющих животных к такой функциональной гипоксии определяется рядом физиологических адаптации, направленных, с одной стороны, на увеличение запасов О2 в организме перед нырянием, а с другой- на более экономное расходование этих запасов во время погружения.
Запасание кислорода при нырянии. Запасы О2 обычно связывают с накоплением его в крови и мышцах, поскольку запас О2 в легких, видимо, невелик. Многие ныряющие животные уходят в воду на выдохе, что более выгодно, поскольку снижает плавучесть и затраты энергии на погружение. Для глубоконыряющих животных это, кроме того, препятствует возникновению «кессонной болезни». Показано, например, что у финвала объем легких составляет всего лишь 3% от объема тела, а у бутылконоса - даже 1%; у человека этот показатель равен 7%. Инструментальные наблюдения пока зали, что тюлени (Halichoerusgrypus, Phocavitulina, Miroungaangustiros, Leptonychotesweddelii) ныряют при заполнении легких на 60-20% от их общего объема.
В результате относительная роль О2, содержащегося в воздухе, заполняющем легкие, в общем его запасании на время погружения оказывается второстепенной (табл. 12). У обыкновенного тюленя из общего запаса О2 в организме (40 мл/кг) только 14 мл/кг приходится на долю легочного запаса, тогда как у человека и собаки соотношение обратное: только 10 мл/кг приходится на внелегочные запасы.
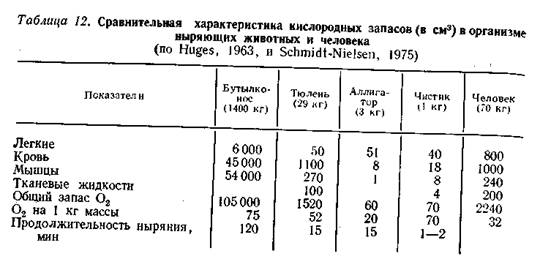
Утки и, видимо, большинство других птиц также ныряют на выдохе. Только для пингвинов (Pygoscelisadeliae, P. papua) доказано ныряние на вдохе; при кратковременности погружения, свойственной этим видам, такая ситуация не грозит им опасным насыщением крови инертными газами. В то же время экспериментально показано, что запас воздуха в дыхательной системе обеспечивает возможность этим птицам продолжать газообмен в легких во время ныряния (Kooyman et al., 1973).
Существенно большее значение имеет для ныряющих животных запасание О2 в крови (табл. 12). Эффективность этого пути определяется общим количеством крови, содержанием гемоглобина, числом и суммарной поверхностью эритроцитов и некоторыми другими гематологическими параметрами, в совокупности определяющими кислородную емкость крови. Отмечено, что эти параметры у хорошо ныряющих животных (китообразные, ластоногие) несколько выше, чем у неныряющих. Это прослеживается и в ряду ныряющих животных: исследование трех видов дельфинов показало, что общее содержание О2 в крови высокоактивной и глубоко ныряющей пелагической белокрылой морской свиньи Phocaenoidesdalli почти в 3 раза выше, чем у прибрежной афалины Tursiopstruncatus, и на 70% больше, чем у пелагического полосатого дельфина Lagenorhynchusobliquidens, отличающегося не очень высокой активностью.
Сходная картина обнаруживается и у птиц: параметры красной крови у ныряющих форм обычно несколько выше, чем у неныряющих. Так, общий объем крови у хохлатых пингвинов в среднем составляет 14,3% от массы тела, у хохлатой чернети-14,6, у красноголового нырка- 15,6, у гаги- 16-17%, тогда как у других водоплавающих около 13%, а у наземных птиц-в среднем около 6% от массы тела. Концентрация гемоглобина у пингвинов колеблется в пределах 17-20 г%, у тупика и кайры этот показатель составляет соответственно 18 и 19,4 г%, а у наземных видов он существенно меньше: у неясыти 7,7%, у домового воробья 10,8, у степного орла 12,6 г% (Коржуев, 1964; Коржуев и др., 1962, 1977).
В результате у птиц-ныряльщиков несколько выше может быть и общая кислородная емкость крови: у пингвинов порядка 20 об. %, у чистиковых 24-26, у домашней утки около 17, а у кур 11,2 об.%; в то же время у голубя кислородная емкость крови составляет 21,2 об.%, что связано с активным полетом (Andersen, 1966).
По признаку сродства гемоглобина к О2 ныряющие животные в большинстве случаев существенно не отличаются от неныряющих. Поскольку дыхание у всех этих животных происходит при нормальном давлении О2, повышение степени сродства не имело бы смысла; при разрядке оксигемоглобина в тканях высокое сродство к О2 скорее, затрудняло бы этот процесс. Относительно высокое сродство гемоглобина к О2 обнаружено у пингвинов, хотя и у них вели чина разрядного напряжения pso (28,8-34,4 мм рт. ст., или 3,8- 4,6 кПа) сопоставима с тем же показателем столь хорошего летуна, как голубь (29,5 мм, или 3,9 кПа). Не исключено, что высокое сродство гемоглобина к О2 выполняет у пингвинов особую функцию, способствуя ускорению реоксигенации крови при коротких появлениях кормящихся птиц на поверхности воды, а относительно выраженный эффект Бора облегчает снабжение тканей О2.
Специальную функциональную направленность имеет запасание О2 в мышцах путем связывания его с миоглобином (табл. 12). Количество миоглобина в мускулатуре ныряющих животных может быть очень большим. У дельфинов такая мускулатура составляет более трети массы тела. И в этом случае важную роль играют экологические особенности различных животных. Установлено, что содержание миоглобина в мышцах дельфина (морская свинья) значительно выше, чем у малоподвижных ламантинов; исключение составляют жевательные мышцы, которые у дюгоня активно работают при погружении в воду (Blessing, 1972). При сравнении утки, лысухи и поганки оказалось, что содержание миоглобина в сердечной мышце выше всего у хорошо ныряющей поганки; у всех видов концентрация миоглобина была выше в левом желудочке, чем в правом, что явно связано с различной нагрузкой (Deshpande,. 1980). У хохлатого пингвина концентрация миоглобина в грудных мышцах составляет в среднем 3700 мг%, а общая обеспеченность организма достигает 10,4 г миоглобина на 1 кг массы тела. У пингвина Адели концентрация миоглобина 2800-3200 мг%, у антарктического пингвина 4200-4600 мг%, тогда как у наземных птиц - порядка 300-400 мг%.
Как уже говорилось, миоглобин обладает сродством к О2 при мерно в 10 раз большим, чем гемоглобин. Поэтому при обычных условиях он легко насыщается О2, транспортируемым с кровью, а в период погружения, когда приток О2 с кровью сокращается или прекращается совсем, отдает связанный ранее О2 ткани, окислительная ферментная система которой способна насыщаться О2 при малых величинах его парциального давления (рис. 19).
Приспособления к экономному расходованию запасов кислорода. Как показывают расчеты, общий запас О2 в легких, крови и. мышцах не в состоянии обеспечить продолжительную остановку дыхания, свойственную ныряющим животным, если он расходуется с такой же скоростью, что и при свободном дыхании. Так, у пингвинов запас О2 перед погружением обеспечивает существование на уровне метаболизма покоя в течение 3 мин, тогда как длительность естественной задержки дыхания при нырянии доходит до 5-7 мин.
Тюлень находится под водой втрое дольше, чем «позволяют» расчетные данные; примерно таково же соотношение расчетных и реальных величин у китов. Все это свидетельствует о том, что расход О2 в организме во время ныряния существенно ниже, чем при нормальном дыхании. Система приспособлений, определяющих этот эффект, имеет очень важное значение для всех ныряющих животных. В основе этих приспособлений лежат сердечно-сосудистые реакции организма.
Одно из наиболее существенных приспособлений к нырянию - урежение сердечного ритма (брадикардия), возникающее с момента погружения в воду. Эта реакция зарегистрирована практически у всех исследованных видов ныряющих животных, включая рептилий (рис. 20, 21).
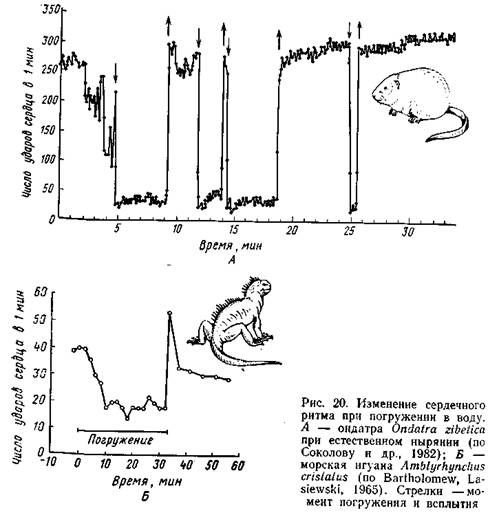
В опытах с тюленями и морскими львами, обученными нырять с укрепленными на теле электродами, было установлено, что при произвольном нырянии частота пульса сначала быстро, а затем замедленно снижалась до 20 ударов в минуту; у тех же животных при насильственном погружении пульс очень быстро (за 2-3 с) падал.до 8 ударов в минуту (Eisner, 1970).
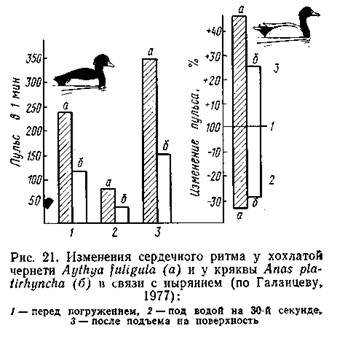
Аналогичные данные получены на птицах. В опыте с домашними утками насильственное погружение в воду вызывало урежение сердечного ритма до 1/4-1/20 от исходного уровня. Телеметрические наблюдения за свободноныряющими красноголовыми нырками и хохлатыми чернетями да ли несколько иные показатели: перед погружением отмечались тахикардия (учащение пульса) и учащенное дыхание, при погружении развивалась брадикардия, после чего частота пульса по степенно восстанавливалась до исходной величины, предшествовавшей всплытию (Butler, Woakes, 1979).
Степень замедления сердечного ритма у разных видов неодинакова. У дельфинов при кратковременных произвольных заныриваниях частота сокращений сердца снижалась вдвое, у тюленя Уэддела (насильственное погружение) - в 3 раза, у морского слона (опыты с погружением головы) - в 4-6 раз, у каланов погружение в воду замедляет сердечный ритм в 2-8 раз.
В результате замедления сердечной деятельности обмен несколько снижается. Так, обыкновенный тюлень, в спокойном со стоянии расходующий около 200 мл О2 в минуту, при нырянии затрачивает всего 50 мл/мин. Показано, что частота пульса меняется Даже у одной особи в зависимости от характера ныряния: у тюле ней Уэддела глубокая брадикардия выражена лишь при длительных погружениях, а при кратковременных ныряниях частота пульса меняется слабо (Kooyman, 1975).
Характерно, что брадикардия возникает очень быстро, задолго до появления кислородной недостаточности. Осуществляется эта реакция рефлекторно, под влиянием сигналов со специфических рецепторов. В частности, указывается на роль механорецепторов, расположенных в дыхательной мускулатуре: в опытах с ондатрами сжатие легких вызывало брадикардию. С другой стороны, наступление брадикардии стимулируется (тоже рефлекторным путем) ощущением влаги в зоне ноздрей и некоторых других частей голо вы. В экспериментах с различными животными смачивание лице вой части головы или только ноздрей стимулировало брадикардию даже без последующего погружения. Регуляция в этом случае осуществляется с помощью сигналов от рецепторов, расположенных в коже головы. При блокировании импульсов с этих рецепторов перерезкой или анестезией лицевого нерва (опыты на тюленях) брадикардия при погружении развивалась значительно медленнее, а при погружении с сохранением дыхания (искусственного) не развивалась вообще (Dykes, 1974).
Ныряющим животным свойственна сниженная чувствительность дыхательного центра головного мозга к некоторому повышению уровня СО2 в крови. К тому же многие из них временно депонируют венозную кровь в расширениях венозных сосудов.
Чрезвычайно существенно приспособление к экономному расходу О2 в виде изменения характера циркуляции крови в организме. Еще в 1940 г. П. Шоландер (Scholander, 1940) обнаружил, что в течение длительного ныряния содержание молочной кислоты в крови тюленей, уток и пингвинов повышается всего в 2-3 раза, но в момент всплытия концентрация ее в 10 раз превышает исходный (до ныряния) уровень. На этом основании он высказал предположение, что образующаяся в мышцах при гликолизе молочная кис лота не поступает в кровь из-за ограничения кровоснабжения мышц во время ныряния; при всплытии циркуляция в мышцах восстанавливается, и молочная кислота в большом количестве переходит в кровяное русло. Последующие исследования подтвердили эту гипотезу. В частности, с помощью ангиографии было показано, что на время ныряния у тюленей периферические кровеносные сосуды сжимаются и кровоток в них прекращается (или сильно замедляется). При этом основная масса мускулатуры практически отключается от снабжения крови кислородом.
Особенно показательны исследования К. Иогансена (Johansen, 1964), который погруженным в воду уткам внутривенно вводил радиоактивный изотоп 86Rb в момент появления брадикардии. Через 2 мин уток забивали и исследовали распределение изотопа в тканях, что служило надежным показателем уровня кровотока в разных частях организма. В результате было установлено, что во время ныряния резко уменьшается (вплоть до полного прекращения) снабжение кровью большей части мускулатуры (кроме мышц головы и пищевода, которые у утки деятельно функционируют при сборе пищи под водой), кишечника, кожи (кроме кожи в области головы), почек, поджелудочной железы. Мозг и сердечная мышца, судя по этим опытам, снабжаются кровью нормально, а в щитовидной железе и надпочечниках отмечено несколько повышенное кровоснабжение.
Практически все данные по сердечно-сосудистым адаптациям к нырянию получены в опытах с насильственным погружением в воду испытуемого животного или только его головы. Немногие исследования на пингвинах (Адели, антарктический) с использованием радиотелеметрической техники позволили получить сведения о реакциях организма при естественном нырянии. Было выяснено, что брадикардия выражена и при естественном погружении пингвинов: частота сердечных сокращений в погруженном состоянии составляла у пингвина Адели 25%, а у антарктического - 70% от исходного (до заныривания) уровня. В отличие от уток у пингвинов отмечено снабжение кровью мускулатуры, хотя и сниженное (Millard et al., 1973). Как указывалось, пингвины ныряют на вдохе, и, по-видимому, во время ныряния продолжается газообмен запасенного воздуха с кровью.
У совершающих короткие (около 1,5 мин) заныривания дельфинов афалин, приученных к условиям опыта в открытом море, так же обнаружена внутримышечная циркуляция крови (Ridaway, Howard, 1979). Вероятно, этот тип адаптации используется лишь при длительном погружении.
Принципы сердечно-сосудистых адаптации в виде возникновения брадикардии и перераспределения тока крови оказываются общими для ныряющих животных. Перераспределение кровотока во время ныряния обеспечивает поддержание нормального, несмотря на брадикардию, артериального давления, а также подачу О2, запасенного в крови, только к органам, активно работающим в погружен ном состоянии. Остальные органы (в первую очередь мускулатура) используют запасы О2 в виде оксимиоглобина, а отчасти переключаются на анаэробные процессы, о чем свидетельствует подчас значительное повышение уровня молочной кислоты как в самих мышцах, так и (после всплытия) в крови.
Биологическая значимость перераспределения кровотока при нырянии подтверждается тем, что в опытах с тюленями выключение сфинктера задней полой вены вело к гибели подопытных животных уже после 4-минутного пребывания под водой (Packer et al., 1969).
Сложная система адаптивных реакций ныряющих животных контролируется центральной нервной системой на уровне продолговатого мозга. В экспериментах на птицах двусторонняя перерезка тройничного нерва снимала рефлекс брадикардии и остановки дыхания при обычных для этих реакций «пусковых механизмах», о которых говорилось выше. Аналогичный эффект вызывала перерезка или холодовой блок блуждающих нервов (Andersen, 1963; Butler, Jones, 1968). Более детальных сведений о механизмах нервной регуляции сердечно-сосудистых реакций при нырянии, так же как о природе рецепторов, не имеется.
Приспособления к гипоксии. Замедление сердечного ритма и изменение характера циркуляции крови.
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 1059; Нарушение авторских прав?; Мы поможем в написании вашей работы!