КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Координация нервных процессов
|
|
|
|
Взаимодействие нейронов, а, следовательно, и нервных процессов, постоянно происходящее в центральной нервной системе, получило название координации нервных процессов. Выделяют следующие разновидности координации нервных процессов в центральной нервной системе.
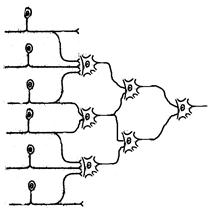 Конвергенция нейронов в центральной нервной системе, заключающаяся в том, что на теле и дендритах какого-то одного вставочного или эфферентного нейрона образуют синапсы множество других нейронов. Общее количество вставочных нейронов в центральной нервной системе в 5 раз превышает таковое для эфферентных. Так, с мотонейронами спинного мозга, иннервирующими определенные скелетные мышцы туловища и конечностей, вступают в синаптический контакт как чувствительные нейроны, несущие информацию от различных рецептивных полей (рецепторов растяжения самих скелетных мышц, рецепторов кожи и некоторых других), так и вставочные нейроны самого спинного мозга и аксоны нейронов головного мозга, проводящие эфферентную информацию к спинному мозгу (нервные волокна т.н. нисходящих проводящих путей). Таким образом, мотонейроны спинного мозга представляют собой общий конечный путь для различных рефлекторных актов.
Конвергенция нейронов в центральной нервной системе, заключающаяся в том, что на теле и дендритах какого-то одного вставочного или эфферентного нейрона образуют синапсы множество других нейронов. Общее количество вставочных нейронов в центральной нервной системе в 5 раз превышает таковое для эфферентных. Так, с мотонейронами спинного мозга, иннервирующими определенные скелетные мышцы туловища и конечностей, вступают в синаптический контакт как чувствительные нейроны, несущие информацию от различных рецептивных полей (рецепторов растяжения самих скелетных мышц, рецепторов кожи и некоторых других), так и вставочные нейроны самого спинного мозга и аксоны нейронов головного мозга, проводящие эфферентную информацию к спинному мозгу (нервные волокна т.н. нисходящих проводящих путей). Таким образом, мотонейроны спинного мозга представляют собой общий конечный путь для различных рефлекторных актов.
Рис. 17. Схема, отражающая конвергенцию нейронов в нервной системе (несколько чувствительных нейронов передают информацию на один вставочный, а несколько вставочных – на один эфферентный нейрон) (по Бабскому Е.Б, 1972).
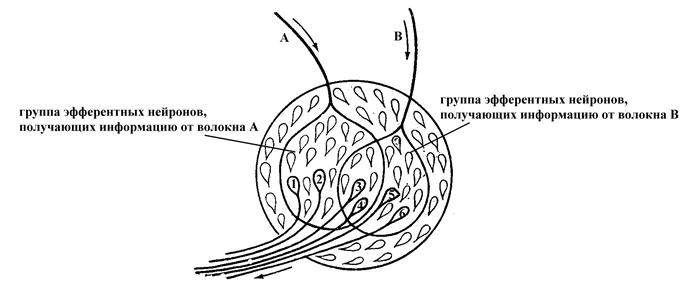 Явление конвергенции нейронов в центральной нервной системе обуславливает, с одной стороны, возможность пространственной суммации информации, поступающей к нейрону через различные его синаптические входы, а с другой, – может служить причиной окклюзии. Сущность окклюзии (закупорки) состоит в том, что если раздражать два соседних чувствительных нервных волокна стимулами определенной силы, то возникающая при этом рефлекторная реакция по силе будет меньше алгебраической суммы рефлекторных ответов, возникающих при раздражении этих волокон по отдельности. Причина такой закупорки заключается в том, что оба эти чувствительные нервные волокна могут вступать в синаптический контакт с одними и теми же эфферентными нейронами. Поэтому при раздражении их по отдельности в возбуждение вовлекается определенное количество нейронов, с которыми вступает в контакт данное чувствительное нервное волокно (в случае примера, представленного на рис. 71, при возбуждении каждого в отдельности афферентного нервного волокна А или В активируются по 4 эфферентных нейрона). Между тем при совместном раздражении обоих чувствительных нервных волокон А и В количество вовлекаемых в рефлекторный ответ эфферентных нейронов будет меньше алгебраической суммы нейронов, задействованных при раздражении каждого в отдельности чувствительного нервного волокна (в частности, в данном примере при совместном возбуждении чувствительных волокон А и В будет задействовано 6, а не 8, эфферентных нейронов), поскольку некоторые эфферентные нейроны являются общими для раздражаемых чувствительных нервных волокон. В связи с отмеченным ответная реакция при совместном раздражении обоих чувствительных нервных волокон (А и В) будет, хоть и больше таковой при раздражении какого-то одного из них, но меньше алгебраической суммы реакций при раздражении афферентных нервных волокон по отдельности.
Явление конвергенции нейронов в центральной нервной системе обуславливает, с одной стороны, возможность пространственной суммации информации, поступающей к нейрону через различные его синаптические входы, а с другой, – может служить причиной окклюзии. Сущность окклюзии (закупорки) состоит в том, что если раздражать два соседних чувствительных нервных волокна стимулами определенной силы, то возникающая при этом рефлекторная реакция по силе будет меньше алгебраической суммы рефлекторных ответов, возникающих при раздражении этих волокон по отдельности. Причина такой закупорки заключается в том, что оба эти чувствительные нервные волокна могут вступать в синаптический контакт с одними и теми же эфферентными нейронами. Поэтому при раздражении их по отдельности в возбуждение вовлекается определенное количество нейронов, с которыми вступает в контакт данное чувствительное нервное волокно (в случае примера, представленного на рис. 71, при возбуждении каждого в отдельности афферентного нервного волокна А или В активируются по 4 эфферентных нейрона). Между тем при совместном раздражении обоих чувствительных нервных волокон А и В количество вовлекаемых в рефлекторный ответ эфферентных нейронов будет меньше алгебраической суммы нейронов, задействованных при раздражении каждого в отдельности чувствительного нервного волокна (в частности, в данном примере при совместном возбуждении чувствительных волокон А и В будет задействовано 6, а не 8, эфферентных нейронов), поскольку некоторые эфферентные нейроны являются общими для раздражаемых чувствительных нервных волокон. В связи с отмеченным ответная реакция при совместном раздражении обоих чувствительных нервных волокон (А и В) будет, хоть и больше таковой при раздражении какого-то одного из них, но меньше алгебраической суммы реакций при раздражении афферентных нервных волокон по отдельности.
|
|
|
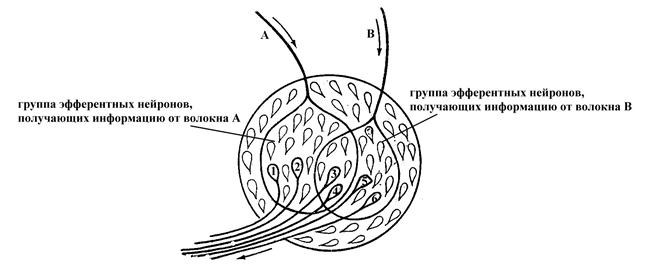
Рис. 18. Схема, раскрывающая причину возникновения окклюзии в нервной системе (по Косицкому Г.И., 1985).
Иррадиация (распространение) возбуждения в центральной нервной системе, обусловлена дивергенцией нервных влияний. Так, аксон одного чувствительного нейрона вступает в синаптический контакт не с одной вставочной или эфферентной нервной клеткой, а с большим количеством других нейронов, зачастую расположенных на разных уровнях центральной нервной системы. Благодаря такой дивергенции нервных влияний возбуждение, возникшее даже в сравнительно небольшой группе афферентных нервных волокон, теоретически могло бы охватить довольно широкие области центральной нервной системы. Вместе с тем, в норме такого генерализованного охвата возбуждением центральной нервной системы при раздражении определенных рецептивных полей не происходит вследствие ограничения процесса иррадиации возбуждения центральным торможением и локализации проведения возбуждения по определенным путям, что является залогом очень тонкого и точного регулирования различных физиологических функций в организме.
|
|
|
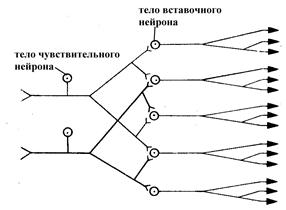 |
Рис. 19. Схема, отражающая пример дивергенции нейронов в нервной системе. Показано, как каждый чувствительный нейрон вступает в контакт с несколькими вставочными, а аксон каждого вставочного, в свою очередь, ветвится и может вступить в контакт с несколькими другими нейронами (вставочными или эфферентными) (по Бабскому Е.Б., 1972).
Возвратное торможение в центральной нервной системе. Сущность данного процесса состоит в том, что определенная нервная клетка (чаще всего эфферентный нейрон), возбуждаясь вследствие поступления к ней информации от других клеток, одновременно возбуждает и специальную тормозную нервную клетку (в спинном мозге тормозными нейронами являются клетки Реншоу), которая, наряду с возможным влиянием на другие нейроны центральной нервной системы, оказывает тормозное воздействие и на сам эфферентный нейрон, возбудивший ее. Благодаря отмеченному возвратному торможению многие эфферентные нейроны могут сами ограничивать свою активность. Так, возбуждаясь в результате поступления информации от других нейронов, они проводят нервные импульсы к исполнительному органу, но одновременно активируют и тормозный нейрон, который образует тормозный синапс на мембране тела самого эфферентного нейрона и ослабляет или полностью прекращает дальнейшую его активность.
|
|
|
 Рис. 20. Схема возвратного торможения мотонейрона в центральной нервной системе (по Бабскому Е.Б., 1972).
Рис. 20. Схема возвратного торможения мотонейрона в центральной нервной системе (по Бабскому Е.Б., 1972).
Реципрокная (сопряженная) иннервация некоторых антагонистичных по функции структур организма (как правило, мышц-антагонистов). Заключается в том, что возбуждение определенных мотонейронов, иннервирующих какие-то скелетные мышцы (например, мышцы-сгибатели), передается не только на сами мышечные волокна, но и на тормозные нейроны, образующие тормозные синапсы на мотонейронах, иннервирующих мышцы антагонистической группы (мышцы-разгибатели). Подобную сопряженную иннервацию скелетных мышц антагонистических групп иногда называют одновременной отрицательной индукцией (возбуждение одной группы мотонейронов через тормозные вставочные нейроны оказывает тормозное влияние на мотонейроны антагонистической группы мышц). Данное явление делает возможным нормальное осуществление многих двигательных актов. Подобная реципрокная иннервация антагонистических по функции органов может, очевидно, осуществляться не только в случае соматической мускулатуры.
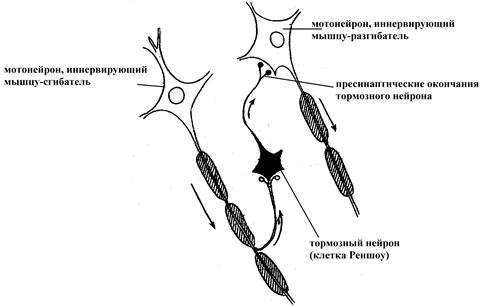 |
Рис. 21. Схема реципрокного торможения мотонейрона в центральной нервной системе (по Бабскому Е.Б., 1972).
Явление последовательной индукции (положительной и отрицательной). Для многих нервных центров, регулирующих различные физиологические функции организма характерно контрастное изменение состояния после прекращения возбуждающего или тормозного раздражения. Так, после прекращения тормозных влияний каких-то нейронов на определенный нервный центр в нем может возникнуть возбуждение (положительная последовательная индукция: возбуждение вслед за торможением). Примером положительной последовательной индукции является возникновение разгибательного рефлекса верхней конечности после продолжительного его торможения вследствие выполнения сгибательного рефлекса. В основе положительной последовательной индукции лежит тот факт, что к нервному центру, который является временно заторможенным по каким-то причинам (например, мотонейроны, иннервирующие мышцы-разгибатели, реципрокно тормозятся при возбуждении мотонейронов, иннервирующих мышцы-сгибатели), продолжает поступать возбуждающая его афферентная информация от рабочих органов (в нашем примере от расслабленной и растянутой мышцы-разгибателя). В связи с отмеченным сразу после прекращения тормозного воздействия на нервный центр в нем возникает возбуждение, инициирующее какой-то новый рефлекс (в нашем случае сокращение мышц-разгибателей). Именно положительная последовательная индукция обуславливает феномен отдачи: быструю смену одного рефлекса другим, противоположным по значению.
|
|
|
В случае если длительное возбуждение нервного центра сопровождается в дальнейшем развитием торможения в нем, говорят об отрицательной последовательной индукции. Одной из причин возникновения отрицательной последовательной индукции может служить длительная следовая гиперполяризация, развивающаяся в нейронах нервного центра после генерации ими нервных импульсов.
Принцип доминанты (сформулирован А.А. Ухтомским). Нервная система состоит из огромного количества нейронов, определенным образом связанных между собой и формирующих нервные центры, ответственные за осуществление какого-то рефлекторного акта или регуляцию определенной физиологической функции. Несмотря на многообразие нервных центров, нервная система работает как единое целое, обеспечивая постоянное нормальное жизнесуществование организма в каждый данный момент. Одним из условий такой слаженной работы нервной системы является определенная взаимосвязь между нервными центрами, осуществляемая по принципу доминанты. Так, для каждого данного момента жизнедеятельности организма в зависимости от его первостепенных потребностей, в центральной нервной системе возникают господствующие очаги возбуждения (доминантные нервные центры), изменяющие и подчиняющие себе работу всех других нервных центров. Примером проявления принципа доминанты является резкое ослабление пищедобывательного рефлекса при нанесении на организм сильного болевого раздражения, сопровождающегося возникновением оборонительной реакции (в данном случае доминантную роль играет нервный центр, обеспечивающий оборонительную реакцию организма, как самую жизненно важную в сложившейся ситуации).
Доминантный очаг возбуждения обладает следующими свойствами:
Ø повышенной возбудимостью
Ø стойкостью возбуждения
Ø способностью к суммации возбуждений
Ø инерцией (способностью к длительному удержанию возбуждения).
|
|
|
|
|
Дата добавления: 2014-11-16; Просмотров: 2125; Нарушение авторских прав?; Мы поможем в написании вашей работы!