КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Экологические системы 1 страница
|
|
|
|
Лекция №5
План:
1. Концепция экосистемы
2. Гомеостаз экосистемы
3. Динамика экосистемы
4. Классификация природных систем биосферы на ландшафтной основе
5. Наземные биомы (экосистемы)
6. Пресноводные экосистемы, морские экосистемы, целостность биосферы как глобальной экосистемы
7. Антропогенные экосистемы
Ключевые слова: экосистема, природные экосистемы, гомеостаз, сукцессия, эндогенный ритм,
Экологическая система, или экосистема, – это «объективно существующая часть природной среды, которая имеет пространственно-территориальные границы и в которой живые (растения, животные и другие организмы) и неживые ее элементы взаимодействуют как единое функциональное целое и связаны между собой обменом веществом и энергией».
Природные экосистемы – это открытые системы: они должны получать и отдавать вещества и энергию. Запасы веществ, усвояемые организмами и прежде всего продуцентами, в природе не безграничны. Если бы эти вещества не использовались многократно, то жизнь на Земле была бы невозможна. Вечный круговорот биогенных компонентов возможен лишь при наличии функционально различных групп организмов, которые осуществляют и поддерживают поток веществ, извлекаемых ими из окружающей среды.
Гомеостаз- способность биологических систем (организма, популяции и экосистем) противостоять изменениям и сохранять равновесие. Исходя из кибернетической природы систем, гомеостатический механизм — это обратная связь. Положительная обратная связь «усиливает отклонение», например, увеличивает чрезмерно популяцию жертвы. Отрицательная обратная связь «уменьшает отклонение», например, ограничивает рост популяции жертвы за счет увеличения численности популяции хищников.
|
|
|
Периодичность внешних условий и проявление внутренних (эндогенных) ритмов организмов, флуктуации популяций достаточно синхронно отражаются в цикличности всего биотического сообщества экосистемы. Изменение численности биоценоза может быть вызвано резко неравномерным выпадением осадков по годам, с периодическим повторением засух, что иллюстрируется повторением массовых размножений животных, например саранчевых.
Энергетика экосистем
Живые организмы, входящие в экосистемы, для своего существования должны постоянно пополнять и расходовать энергию. Растения, как известно, способны запасать энергию в химических связях в процессе фотосинтеза или хемосинтеза. При фотосинтезе связывается только энергия с определенными длинами волн -380-710 нм. Эту энергию называют фотосинтетически активной радиацией (ФАР). Она по длинам волн близка к видимой части спектра. На эту радиацию обычно приходится около 40% общей солнечной радиации, достигающей земной поверхности. Остальная часть спектра относится либо к более короткой (ультрафиолетовой), либо к более длинной (инфракрасной) радиации. С последней обычно связан тепловой эффект.
Растения в процессе фотосинтеза связывают лишь небольшую часть солнечной радиации. Даже по отношению к фотосинтетически активной - это в среднем для земного шара менее 1%. Только наиболее продуктивные экосистемы, такие как плантации сахарного тростника, тропические леса, посевы кукурузы, в оптимальных условиях могут связывать до 3-5% ФАР. В опытах с кондиционированными условиями по всем факторам среды за короткие периоды времени удавалось достичь эффективности фотосинтеза по усвоению солнечной энергии порядка 8-10% ФАР.
Растения являются первичными поставщиками энергии для всех других организмов в цепях питания. Существуют определенные закономерности перехода энергии с одного трофического уровня на другой вместе с потребляемой пищей. Основная часть энергии, усвоенной консументом с пищей, расходуется на его жизнеобеспечение (движение, поддержание температуры тела и т. п.). Эту часть энергии рассматривают как траты на дыхание, с которым в конечном счете связаны все возможности ее высвобождения из химических связей органического вещества.
|
|
|
Часть энергии переходит в тело организма-потребителя вместе с увеличивающейся массой (приростом, продукцией). Некоторая доля пищи, а вместе с ней и энергия не усваиваются организмом. Они выводятся в окружающую среду вместе с продуктами жизнедеятельности (экскрементами). В последующем эта энергия высвобождается другими организмами, которые потребляют продукты выделения.
Баланс пищи и энергии для отдельного животного организма можно, таким образом, представить в виде следующего уравнения:
Эп = Эд +Эпр +Эп.в,
где Эп - энергия потребленной пищи, Эд - энергия дыхания или обеспечения жизнедеятельности организма, включая движение, поддержание температуры тела, сердцебиение и т. п., Эпр - энергия прироста (запасенная в теле организма-потребителя), Эп.в - энергия продуктов выделения (в основном экскрементов).
Количество энергии, расходуемой организмами на различные цели, неоднозначно. В периоды интенсивной жизнедеятельности взрослого организма в теле его может совершенно не фиксироваться энергия. Наоборот, траты ее в ряде случаев превышают поступление (организм теряет вес). В то же время в периоды интенсивного роста организмов, особенно в периоды размножения (беременности), в теле фиксируется значительное количество энергии.
Выделение энергии с экскрементами у плотоядных животных (например, хищников) невелико, у травоядных оно более значительно, а гусеницы некоторых насекомых, питающиеся растениями, выделяют с экскрементами до 70% энергии. Однако при всем разнообразии расходов энергии в среднем максимальны траты на дыхание, которые в сумме с неусвоенной пищей составляют около 90% от потребленной. Поэтому переход энергии с одного трофического уровня на другой в среднем принимается близким к 10% от энергии, потребленной с пищей. Эта закономерность рассматривается обычно как «правило десяти процентов».
|
|
|
Данное правило надо оценивать как относительное, ориентировочное. Вместе с тем из него следует, что цепь питания имеет ограниченное количество уровней, обычно не более 4—5. Пройдя через них, практически вся энергия оказывается рассеянной.
Закономерности потока и рассеивания энергии имеют важные в практическом отношении следствия. Во-первых, с энергетической точки зрения крайне нецелесообразно потребление животной продукции, особенно с высоких уровней цепей питания. Образование этой продукции связано с большими потерями (рассеиванием) энергии. Особенно велики потери энергии при переходе с первого трофического уровня на второй, от растений к травоядным животным.
Часто в экологической литературе рассматривается в качестве примера цепь питания: люцерна-телята-мальчик. Показано, что если бы мальчик весом 48 кг питался только телятиной, то за год ему потребовалось бы для обеспечения жизнедеятельности 4,5 теленка, для питания которых, в свою очередь, необходим урожай люцерны с площади 4 га весом 8211 кг. Такова энергетическая цена животной пищи.
Во-вторых, чтобы сократить вероятность дефицита продуктов питания для интенсивно возрастающей численности населения (по закономерности, близкой к экспоненте), надо, чтобы в рационе людей больший удельный вес занимала растительная пища. Энергетически идеально - вегетарианство.
В-третьих, для увеличения КПД использования пищи при получении животноводческой продукции в условиях культурного хозяйства очень важно уменьшить основную статью нерационального расходования энергии - ее траты на дыхание. Это возможно за счет поддержания оптимального температурного режима в животноводческих помещениях, ограничения подвижности животных и, естественно, сбалансированности кормового рациона по различным элементам питания, а также применения различных биотехнических приемов (умеренные добавки стимуляторов роста, веществ, способствующих улучшению аппетита и т. п.).
|
|
|
Споры о допустимо возможной численности населения с точки зрения обеспечения питанием в значительной мере относительны, если они не учитывают, какой в среднем удельный вес в рационе отводится животной и растительной пище. Если исходить из рациона питания зажиточной части населения, потребляющей мяса 80-100 кг в год на одного человека, то явно невозможно обеспечение таким рационом современной численности населения Земли (около 6 млрд. человек). Если же исходить из необходимости обеспечения минимальных потребностей жизнедеятельности организма, при настоящем производстве продуктов питания возможно исключить голод и, кроме того, прокормить на 3-4 миллиарда населения больше современного. Для этого требует решения вопрос более сбалансированного распределения продуктов питания. Переход на вегетарианство и тем более расширение ассортимента растений, используемых в пищу, может обеспечить жизнедеятельность (с энергетической точки зрения) численности населения в 2-3 раза больше современной. Ясно, однако, что при этом останутся нерешенными многие медико-биологические проблемы здоровья и долголетия, а также допустимые пределы антропогенных нагрузок на экосистемы и биосферу в целом.
Продуктивность и биомасса экосистем
Одно из важнейших свойств организмов, их популяций и экосистем в целом - способность создавать органическое вещество, которое называют продукцией. Образование продукции в единицу времени (час, сутки, год) на единице площади (метры квадратные, гектар) или объема (в водных экосистемах) характеризует продуктивность экосистем. Продукция и продуктивность могут определяться для экосистем в целом или для отдельных групп организмов (растений, животных, микроорганизмов) или видов.
Продукцию растений называют первичной, а животных - вторичной. Наряду с продукцией различают биомассу организма, групп организмов или экосистем в целом. Под ней понимают всю живую органическую массу, которая содержится в экосистеме или ее элементах вне зависимости от того, за какой период она образовалась и накопилась. Биомасса и продукция (продуктивность) обычно выражаются через абсолютно сухой вес.
Нетрудно понять, что величина биомассы экосистем или их звеньев во многом зависит не столько от их продуктивности, сколько от продолжительности жизни организмов и экосистем в целом. Например, большая биомасса характерна для лесных экосистем: в тропических лесах она достигает 800-1000 т/га, в лесах умеренной зоны -300-400 т/га, а в травянистых сообществах обычно не выходит за пределы 3-5 т/га. В то же время лесные и травянистые (например, луговые) экосистемы в сходных условиях существования по продуктивности могут мало различаться или различаются в сторону большей продуктивности как лесных, так и травянистых сообществ.
Для экосистем, представленных однолетними организмами, их годичная продуктивность и биомасса практически совпадают. Для древесных сообществ они резко различаются. Вообще соотношение биомассы и годовой продукции экосистем можно выразить формулой:
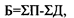
где Б - биомасса в данный момент времени, П - годовая продукция, Д - дыхание. Под последним применительно к экосистемам понимается вся сумма живого вещества, отчуждаемого на процессы разложения в результате гибели целых организмов (отпад) или их частей - сучьев, коры, листьев, наружных покровов (опад) и потребления гетеротрофами.
Экологические параметры продуктивности. Продукция и биомасса экосистем - это не только ресурс, используемый в пищу или в качестве различных видов сырья (техническое, топливо и т. п.). От этих показателей в прямой зависимости находится средообразующая и средостабилизирующая роль экосистем. Так, с продуктивностью растений и их сообществ тесно связана интенсивность поглощения углекислого газа и выделения кислорода. Для образования одной тонны растительной продукции (абсолютно сухой вес) обычно поглощается 1,5-1,8 т углекислого газа и выделяется 1,2-1,4 т кислорода. Биомасса, в том числе и мертвое органическое вещество, являются основными резервуарами концентрации углерода. На суше это практически единственный фактор вывода углекислого газа из процессов круговорота на длительное время. Часть этого органического вещества и вовсе исключается из круговорота или, как отмечал В. И. Вернадский, «уходит в геологию» (торф, уголь, нефть и т. п.).
Чаще всего в гумидных (влажных) районах фактором, прерывающим круговорот, выступает недостаток кислорода и кислая среда. Здесь основными очагами накопления органики являются болота. На дне глубоких водоемов захоронение органического вещества также обусловливается недостатком кислорода или избытком ядовитых веществ (например, сероводорода). В крайне сухих (аридных) условиях круговорот прерывается чаще всего недостатком влаги.
В связи с тем, что дождевые тропические леса характеризуются максимальной продуктивностью (до 20-25 т/га/год) и биомассой (до 700-1000 т/га), их рассматривают как основные аккумуляторы углерода и обогащения атмосферы кислородом, называя «легкими планеты». В северных лесах, как известно, продуктивность (6-10 т/га/год) и биомасса (300-400 т/га) значительно ниже. Однако на этом основании северным лесам никак нельзя отводить менее значительную роль в положительном балансе кислорода и углекислоты. Наоборот, их роль в этом отношении часто более значительна. Эти вопросы рассмотрены во второй части учебника.
Есть и другие экологические аспекты продуктивности и биомассы экосистем. В частности, чем больше биомасса, тем сильнее ее контакт с окружающей средой и тем значительнее такие средоохранные свойства, как очистка воздуха от пыли и химических агентов, регулирование влагооборотов, гашение шумовых воздействий и т. п.
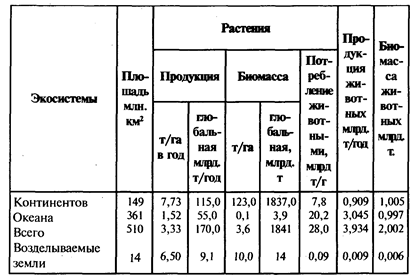
Продуктивность различных экосистем биосферы. До недавнего времени принималось за аксиому, что основной объем первичной продукции образуется в морях и океанах, на долю которых приходится около 70% поверхности земного шара. Однако по последним данным, полученным в основном в результате осуществления Международной биологической программы (МБП), которая проводилась в 1964-1974 гг., было установлено, что основная масса первичной продукции образуется в экосистемах суши (около 115 млрд. тонн в год) и только около 55 млрд. тонн в год - в экосистемах океана (табл. 2). Дело в том, что внутренние воды океана, расположенные за пределами прибрежной (шельфовой) зоны, по продуктивности близки к пустыням наземных экосистем (10-120 г/м2 за год первичной продукции). Для сравнения отметим, что продуктивность лесов тайги составляет в среднем около 700-800, а влажных тропических лесов - 2000-2200 г/м2 за год.
Второй вопрос, на который важно получить ответ: какие же экосистемы в пределах океана и суши являются наиболее продуктивными?
В. И. Вернадский в свое время выделил очаги наибольшей концентрации жизни, назвав их пленками и сгущениями живого вещества. Под пленками живого вещества понимается его повышенное количество на больших пространствах. В океане обычно выделяют две пленки: поверхностную, или планктонную, и донную, или бентосную. Мощность поверхностной пленки обусловливается в основном эуфотической зоной, то есть тем слоем воды, в котором возможен фотосинтез. Она колеблется от нескольких десятков и сотен метров (в чистых водах) до нескольких сантиметров (в загрязненных водах). Донная пленка образована в основном гетеротрофными экосистемами, и поэтому ее продукция представлена вторичной, а количество ее зависит в основном от поступления органического вещества с поверхностной пленки.
В наземных экосистемах также выделяют две пленки живого вещества. Приземная, заключенная между поверхностью почвы и верхней границей растительного покрова, имеет толщину от нескольких сантиметров (пустыни, тундры, болота и др.) до нескольких десятков метров (леса). Вторая пленка - почвенная. Эта пленка наиболее насыщена жизнью. На 1 м2 почвенного слоя насчитывают миллионы насекомых, десятки и сотни дождевых червей и сотни миллионов микроорганизмов. Толщина данной пленки находится в прямой зависимости от мощности почвенного слоя и его богатства гумусом. В тундрах и пустынях это несколько сантиметров, на черноземах, особенно тучных, - до 2-3 метров.
Повышенные концентрации живого вещества в биосфере обычно приурочены к условиям так называемого «краевого эффекта», или экотонов. Такой эффект возникает на стыках сред жизни или различных экосистем. В приведенных примерах для водных экосистем поверхностная пленка - это зона контакта атмосферы и водной среды, донная - водной толщи и донных отложений, почвенная - атмосферы и литосферы.
Таблица 2
Продуктивность и биомасса экосистем материков и океанов земного шара (Уиттекер, 1980)
Примером повышенной продуктивности на стыках экосистем могут служить переходные экосистемы между лесом и полем («опушечный эффект»), а в водных средах - экосистемы, возникающие в эстуариях рек (места впадения их в моря, океаны и озера и т. п.).
Этими же закономерностями во многом обусловливаются упоминавшиеся выше локальные сгущения больших масс живого вещества (наиболее высокопродуктивные экосистемы).
Обычно в океане выделяют следующие сгущения жизни:
1. Прибрежные. Они располагаются на контакте водной и наземно-воздушной среды. Особенно высокопродуктивны экосистемы эстуариев. Протяженность этих сгущений тем значительнее, чем больше вынос реками органических и минеральных веществ с суши.
2. Коралловые рифы. Высокая продуктивность этих экосистем связана прежде всего с благоприятным температурным режимом, фильтрационным типом питания многих организмов, видовым богатством сообществ, симбиотическими связями и другими факторами. 3. Саргассовые сгущения. Создаются большими массами плавающих водорослей, чаще всего саргассовых (в Саргассовом море) и филлофорных (в Черном море). 4. Апвеллинговые. Эти сгущения приурочены к районам океана, где имеет место восходящее движение водных масс от дна к поверхности (апвеллинг). Они несут много донных органических и минеральных отложений и в результате активного перемешивания хорошо обеспечены кислородом. Эти высокопродуктивные экосистемы являются одним из основных районов промысла рыб и других морепродуктов. 5. Рифтовые глубоководные (абиссальные) сгущения. Эти экосистемы были открыты только в 70-х годах настоящего столетия. Они уникальны по своей природе: существуют на больших глубинах (2-3 тыс. метров). Первичная продукция в них образуется только в результате процессов хемосинтеза за счет высвобождения энергии из сернистых соединений, поступающих из разломов дна (рифтов). Высокая продуктивность здесь обязана прежде всего благоприятным температурным условиям, поскольку разломы одновременно являются очагами выхода из недр подогретых (термальных) вод. Это единственные экосистемы, не использующие солнечную энергию. Они живут за счет энергии недр Земли.
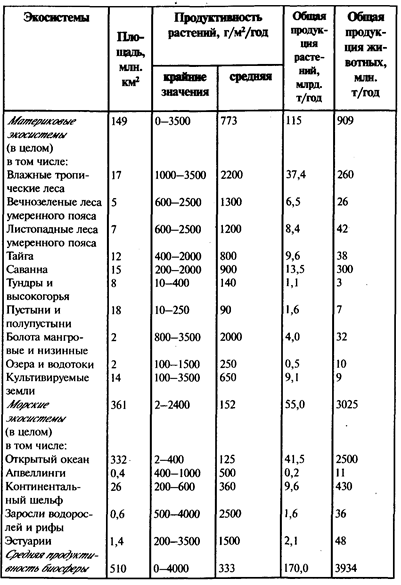
На суше к наиболее высокопродуктивным экосистемам (сгущениям живого вещества) относят: 1 ) экосистемы берегов морей и океанов в районах, хорошо обеспеченных теплом; 2) экосистемы пойм, периодически заливаемые водами рек, которые откладывают ил, а вместе с ним органические и биогенные вещества, 3) экосистемы небольших внутренних водоемов, богатые питательными веществами, а также 4) экосистемы тропических лесов. Продуктивность других экосистем видна из табл.3. Выше мы уже отмечали, что человек должен стремиться сохранить высокопродуктивные экосистемы - этот мощнейший каркас биосферы. Его разрушение связано с наиболее значительными отрицательными последствиями для всей биосферы.
Что касается вторичной (животной) продукции, то она заметно выше в океане, чем в наземных экосистемах. Это связано с тем, что на суше в звено консументов (травоядных) в среднем включается лишь около 10% первичной продукции, а в океане - до 50%. Поэтому, несмотря на более низкую первичную продуктивность океана, чем суши, по массе вторичной продукции эти экосистемы примерно равны (см. табл. 2).
В наземных экосистемах основную продукцию (до 50%) и особенно биомассу (около 90%) дают лесные экосистемы. Вместе с тем основная масса этой продукции поступает сразу в звено деструкторов и редуцентов. Для таких экосистем характерно преобладание детритных (за счет мертвого органического вещества) цепей питания. В травянистых экосистемах (луга, степи, прерии, саванны), как и в океане, значительно большая часть первичной продукции прижизненно отчуждается фитофагами (травоядными животными). Такие цепи носят название пастбищных или цепей выедання.
Экологические пирамиды
Если количество энергии, продукции, биомасс или численности организмов на каждом трофическом уровне изображать в виде прямоугольников в одном и том же масштабе, то их распределение будет иметь вид пирамид.
Таблица 3
Продуктивность основных экосистем земного шара (по Н. Ф. Реймерсу, 1990)
Правило пирамид энергии можно сформулировать следующим образом: количество энергии, содержащейся в организмах на любом последующем трофическом уровне цепи питания, меньше ее значений на предыдущем уровне (рис. 4а).
Рис. 4
Пирамиды энергии и продукции для экосистем суши и океана - а и биомасс для экосистем океана — б
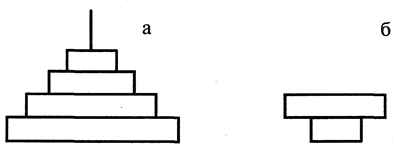
Количество продукции, образующейся в единицу времени на разных трофических уровнях, подчиняется тому же правилу, которое характерно для энергии: на каждом последующем уровне количество продукции меньше, чем на предыдущем (рис. 4а). Более того, суммарное количество вторичной продукции (как и содержащейся в ней энергии), образующейся на разных трофических уровнях, меньше первичной продукции. Эта закономерность абсолютна и легко объясняется исходя из правила передачи энергии в цепях питания. Следует также иметь в виду, что различия в количестве энергии, содержащейся в единице веса (объема) отдельных видов продукции, невелики: 1 г (сухой вес) растительной и животной продукции содержит чаще всего от 3 до 5 калорий энергии.
Пирамиды биомасс сходны с таковыми для энергии и продукции, но только для сухопутных экосистем. Для водных экосистем закономерности соотношения биомасс на различных трофических уровнях имеют свою специфику. Здесь пирамида биомасс как бы перевернута (рис. 4б), то есть биомасса животных, потребляющих растительную продукцию, больше биомассы растительных организмов. Причина этого - резкие различия в продолжительности жизни организмов сравниваемых уровней. Первый уровень (продуценты) представлен в основном фитопланктоном с крайне коротким периодом жизни (несколько дней или часов), второй -более долгоживущими организмами - зоопланктоном или другими животными, питающимися фитопланктоном и зоопланктоном (рыбы, моллюски, киты и т. п.). Они накапливают биомассу годами и десятилетиями.
Пирамида чисел свидетельствует, что количество организмов, как правило, уменьшается от основания к вершине. Это правило не абсолютно и применимо в основном к цепям питания, не включающим редуцентов. Примером может служить пищевая цепь: насекомые и их личинки - насекомоядные животные - хищники.
Динамика и развитие экосистем. Сукцессии
Любая экосистема, приспосабливаясь к изменениям внешней среды, находится в состоянии динамики. Эта динамика может касаться как отдельных звеньев экосистем (организмов, популяций, трофических групп), так и системы в целом. При этом динамика может быть связана, с одной стороны, с адаптациями к факторам, которые являются внешними по отношению к экосистеме, а с другой - к факторам, которые создает и изменяет сама экосистема.
Самый простой тип динамики - суточный. Он связан с изменениями в фотосинтезе и транспирации (испарении воды) растений. В еще большей мере эти изменения связаны с поведением животного населения. Одни из них более активны днем, другие - в сумерки, третьи - ночью. Аналогичные примеры можно привести по отношению к сезонным явлениям, с которыми еще больше связана активность жизнедеятельности организмов.
Не остаются неизменными экосистемы и в многолетнем ряду. Если в качестве примера взять лес или луг, то не трудно заметить, что в разные годы этим экосистемам свойственны свои особенности. В одни годы мы можем наблюдать увеличение численности одних видов (на лугах, например, бывают «клеверные годы», годы с резким увеличением злаков и других видов или групп видов). Из этого следует, что каждый вид индивидуален по своим требованиям к среде, и ее изменения для одних видов благоприятны, а на другие, наоборот, оказывают угнетающее влияние. Сказывается также и периодичность в интенсивности размножения.
Эти изменения в одних случаях могут в какой-то мере повторяться, в других же имеют однонаправленный, поступательный характер и обусловливают развитие экосистемы в определенном направлении.
Периодически повторяющуюся динамику называют циклическими изменениями или флуктуациями, а направленную динамику именуют поступательной или развитием экосистем. Для последнего вида динамики характерным является либо внедрение в экосистемы новых видов, либо смена одних видов другими. В конечном же счете происходят смены биоценозов и экосистем в целом. Этот процесс называют сукцессией (лат. сукцессио - преемственность, наследование). Различают обычно первичные и вторичные сукцессии.
Первичные сукцессии. Под первичной обычно понимается сукцессия, развитие которой начинается на изначально безжизненном субстрате. Ход первичной сукцессии рассмотрим на примере наземных экосистем. Если взять участки земной поверхности, например заброшенные песчаные карьеры, в различных географических районах (в лесной, степной зонах либо среди тропических лесов и т. п.), то для всех этих объектов будут характерны как общие, так и специфические изменения в экосистемах.
В качестве общих закономерностей будет иметь место заселение живыми организмами, увеличение их видового разнообразия, постепенное обогащение почв органическим веществом, возрастание их плодородия, усиление связей между различными видами или трофическими группами организмов, уменьшение числа свободных экологических ниш, постепенное формирование все более сложных биоценозов и экосистем, повышение их продуктивности. Более мелкие виды организмов, особенно растительных, при этом, как правило, сменяются более крупными, интенсифицируются процессы круговорота веществ и т. п. В каждом случае при этом можно выделить последовательные стадии сукцессий, под которыми понимается смена одних экосистем другими, а сукцессионные ряды заканчиваются относительно мало изменяющимися экосистемами. Их называют климаксными (греч. климакс - лестница), коренными или узловыми.
Специфические закономерности сукцессий заключаются прежде всего в том, что каждой из них, как и каждой стадии, присущ тот набор видов, которые, во-первых, характерны для данного региона, а во-вторых, наиболее приспособлены к той или иной стадии развития сукцессионного ряда. Различными будут и завершающие (климаксные) сообщества (экосистемы).
Американский эколог Клементс, наиболее полно разработавший учение о сукцессиях, считает, что в любом обширном географическом районе, который по масштабам можно примерно приравнять к природной зоне (лесная, степная, пустынная и т. п.), каждый ряд завершается одной и той же климаксной экосистемой (моноклимаксом). Такой климакс был назван климатическим. Это, однако, не значит, что для любого участка географической зоны (моноклимакса) характерен один и тот же набор видов. Видовой состав климаксных экосистем может существенно различаться. Общим является лишь то, что эти экосистемы объединяет сходство видов-эдификаторов, то есть тех, которые в наибольшей мере создают среду обитания. Например, для степных экосистем эдификаторами являются плотнокустовые злаки (ковыль и типчак). Для тропических лесов в качестве эдификаторов выступает большое количество древесных видов, создающих сильное затенение для других видов своим пологом.
Для лесной зоны северных и срединных регионов Евразии основными эдификаторами выступают ель или пихта. Из набора всех древесных видов они в наибольшей степени изменяют условия место-произрастания: сильно затеняют подпологовое пространство, создают кислую среду почв и обусловливают процессы их оподзоливания (растворение и вымывание из приповерхностного слоя практически всех минералов, кроме кварца). С этими эдификаторами уживаются только те древесные виды, которые не отстают от них в росте и способны первыми захватить пространство. При сочетании таких условий возможно формирование климаксных смешанных елово-лиственных (пихтово-лиственных), чаще всего с березой и осиной, лесов. Последнее наиболее характерно для зоны смешанных лесов. Для таежной (более северной) зоны более типичны климаксные леса с явным преобладанием только эдификаторов (ель, пихта).
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 1013; Нарушение авторских прав?; Мы поможем в написании вашей работы!