КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Вирусы гриппа типа а
|
|
|
|
Вирус гриппа человека выделен в 1933 г. Смитом, Эндрюсом и Лейдлоу путем заражения хорьков смывами носоглотки больного гриппом Вильсона Смита (отсюда название первого штамма WS).
Морфология. Вирус имеет сферическую форму, диаметр вирусной частицы 80—120 нм (см. рис. 7, а). В препаратах свежевыделенных от больного вирусов встречаются нитевидные формы значительной длины. Вирус содержит липопротеидную оболочку, покрытую шипи-ками длиной около 10 нм. Шипики образованы двумя гликопротеидами — гемагглютинином и нейраминидазой. Шип гемагглютинина образован тремя молекулами белка, шип нейраминидазы образован четырьмя молекулами белка (рис. 43).
Внутренним компонентом вирионов является нуклео-капсид спиральной симметрии с диаметром около 9 нм. Он уложен в виде двойной спирали с диаметром 50—60 нм с односпиральной петлей на конце.
В соответствии с 8 фрагментами генома имеется 8 фрагментов нуклеокапсида. В составе вирионов между фрагментами существует слабая связь.
Химический состав и физико-химические свойства. Вирус содержит по массе 1—2% РНК, 50—70% белков, 18—37% липидов, 5—9% углеводов. Липиды содержатся в липопротеидной оболочке, углеводы — в составе гликопротеидов. И липиды и углеводы имеют клеточное происхождение.
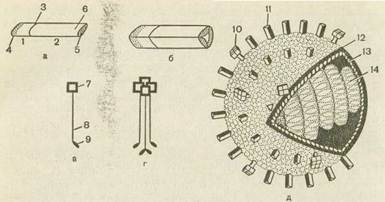
Рис. 43. Строение вириона гриппа (схема). а—мономер гемагглютинина: 1—малая субъединица гемагглютинина; 2— большая субъединица гемагглютинина; 3— место нарезания гемагглютинина-. предшественника; 4— С-конец малой гемагглютинирующей субъединицы, погруженный в липиды; 5— N-конец большой гемагглютинирующей субъединицы, взаимодействующий с рецепторами клетки; б— антигенная детерминанта;
6 — тример гемагглютинина; в — мономер нейраминидазы; 7— гидрофильный участок нейраминидазы; 8 — стержень нейраминидазы; 9 — гидрофобный участок нейраминидазы; г — тетрамер нейраминидазы; 10 — нейраминидаза в составе вириона; 11—гемагглютинин в составе вириона; 12—липидный бислой наружной оболочки вириона; 13 — слой матриксного белка; 14 — рибонуклеопротеид.
|
|
|
Молекулярная масса вирусной частицы 250- 106, плавучая плотность в сахарозе 1,19 г/см3. Фрагменты нуклеокапсида имеют длину от 50 до 130 нм; плавучая плотность нуклеокапсида в хлориде цезия 1,34—1,35 г/см3.
Геном. Геном представлен 8 фрагментами однонитчатой линейной «минус-нитевой» РНК. Молекулярная масса отдельных фрагментов от 0,2- 106 до 1- 106, общая молекулярная масса 5- 106. Каждый фрагмент, за исключением 7-го и 8-го, кодирует один белок, фрагменты 7 и 8 кодируют по два белка с уникальными последовательностями аминокислот, благодаря сплайсингу и сдвигу рамки при трансляции.
Определена первичная структура всех генов вируса гриппа.
Белки, антигены. В составе вируса гриппа содержится 7 белков.
Белки р[, Pa, Рз названы по первой букве слова «по-лимераза», поскольку предполагалось, что все три являют
ся субъединицами РНК-полимеразы. На самом деле, РНК-полимеразой является белок Pi. Белок Рз обладает свойствами эндонуклеазы, «откусывая» «шапочку» у клеточных иРНК вместе с 10—13 прилегающими нуклеотидами и перебрасывая ее на 5/-кoнeц вирусного транскрипта. Функции белка Рг неизвестны. Белки Pi и Рз обладают основными свойствами, а белок ?2 — кислыми, поэтому их еще называют PBi (Pi), PB2 (Рз) и РА (Рз). Все три белка находятся в составе нуклеокапсида и образуют комплекс, интимно связанный с геномом.
Мономер гемагглютинина имеет форму палочки длиной 14 нм и диаметром 4 нм, которая одним концом (С-конец легкой цепи) погружена в липидный бислой (рис. 44). Субъединица гемагглютинина представляет собой тример.
|
|
|
Гемагглютинин синтезируется в виде предшественника, который нарезается протеолитическими ферментами на две субъединицы, тяжелую цепь (НА1) и легкую (НА2) с молекулярными массами примерно 50 000 и 25 000, связанные дисульфидными связями.
С помощью рентгеноструктурного анализа кристаллизованного НА, сделанного Д. Скейлом с соавт., предложена его пространственная модель. Согласно этой модели в тримере НА обнаруживаются два структурных участка — стебель и глобула. Глобула содержит антигенный и рецепторный участки (см. рис. 44). Она состоит только из НА1, а стебель — из обеих субъединиц (НА1 и НА2).
Обнаружено четыре предполагаемые зоны антигенов А, В, С, D. Зона антигена А является петлей, которая образуется между тримерами. Эти зоны соответствуют четырем антигенным детерминантам. Все они связаны с тяжелой цепью гемагглютинина. Две из них являются последовательными, а две образуются при взаимодействии мономеров. Третья обусловлена третичной структурой, четвертая — четвертичной структурой и формируется путем взаимодействия трех мономеров (см. рис. 44). Наиболее активными антигенными детерминантами являются 1-я и 2-я.
Гемагглютинин является основным специфическим антигеном вируса, определяющим, наряду с нейрамини-дазой, подтип вируса и вызывающий образование про-тективных антител. Свойство гемагглютинина ,к антигенной изменчивости обусловлено двумя генетическими процессами — дрейфом и шифтом. В результате' дрейфа происходят незначительные изменения гена НА, обусловленные точечными мутациями, а - антигенные свойства
|
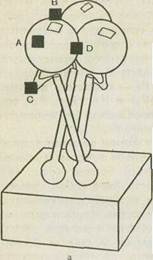
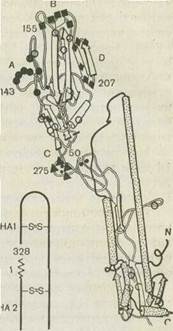
Рис. 44. Гемагглютинин вируса гриппа.
а — тример с четырьмя антигенными детерминантами (черные квадратики^ и участком взаимодействия с клеточным рецептором (белый квадратик — рецепторный карман); б — структура мономера по данным рентгеноструктур-ного анализа. Цифры обозначают порядковый номер аминокислот, кружки, квадраты и треугольники—4 антигенные детерминанты (А, В, С и D), 1—' место нарезания предшественника (N-конец. малой субъединицы), S-S-дисуль" фидные связи.
меняются незначительно, но при продолжительном периоде циркуляции вируса под влиянием коллективного иммунитета селекционируются варианты, значительно отличающиеся по антигенным свойствам от прототипного штамма. Шифт возникает в результате полной смены гена и обусловлен их пересортировкой при одновременной репродукции в клетке двух вирусов гриппа.
|
|
|
Вирус агглютинирует эритроциты кур, морских свинок, человека и многих других видов животных. При низких значениях рН (5,0—5,5) вирус вызывает гемолиз эритроцитов.
Нейраминидаза является ферментом, катализирующим отщепление сиаловой кислоты от субстрата. Мономер нейраминидазы под электронным микроскопом имеет вид
барабанной палочки и состоит из головки, обращенной кнаружи, размером 4Х4Х4 нм и ножки длиной 10 нм. Соединение двух мономеров происходит за счет дисуль-фидных связей, а двух димеров в тетрамере — за счет межмолекулярных связей. Активные центры и антигенные детерминанты находятся на головках мономеров.
Нейраминидаза может изменяться независимо от гемагглютинина, в основе антигенной изменчивости также лежат процессы дрейфа и шифта. Антитела против нейраминидазы не оказывают такого защитного действия, как антитела против гемагглютинина, однако, они частично нейтрализуют вирус и ослабляют инфекционный процесс.
Нуклеопротеид (NP) — основной внутренний белок вируса, формирующий субъединицы капсида. Он интимно связан с геномом в течение всего периода репродукции вируса, но не препятствует экспрессии генома. Этот белок является типоспецифическим антигеном, общим для всех вирусов типа А. Антитела к NP не оказывают защитного действия и применяются с целью диагностики инфекции.
Матриксный белок — самый низкомолекулярный белок в составе вириона, локализованный на внутренней поверхности липопротеидной мембраны вируса. m|-белок играет роль медиатора при сборке вирусной частицы. Mi-белок является стабилизирующим фактором, при его отсутствии или уменьшении концентрации вирусная частица становится хрупкой и быстро распадается.
Матриксный белок, как и NP, является типоспецифическим антигеном, общим для всех вирусов типа А. Белок М2, который кодируется тем же седьмым геном, является неструктурным и локализован в клеточной оболочке.
|
|
|
Белки ns] и NSz — неструктурные вирусные белки, которые хотя и кодируются одним (восьмым) геном, имеют уникальные последовательности аминокислот благодаря трансляции со сдвигом рамки. Белок ns) синтезируется на самых ранних этапах репродукции. В ядре и цитоплазме он накапливается в больших количествах и образует включения.
Чувствительность к физическим и химическим факторам. Вирус гриппа относительно стабилен и может сохраняться при температуре 4° С в течение недели. Инфекционная активность сохраняется продолжительное время при хранении при температуре —70° С. Прогревание при температуре 50—60° С уничтожает его активность
в течение нескольких минут. Вирусы чувствительны к эфиру, детергентам. При низких значениях рН (3,0 и ниже) инфекционная активность теряется.
Репродукция. Вирус адсорбируется на клеточных рецепторах, содержащих сиаловую кислоту (гликопро-теидах или ганглиозидах). Проникновение вируса в клетку Осуществляется обычно по механизму рецепторного Эндоцитоза с последующим слиянием вирусной мембраны со стенкой вакуоли и выходом нуклеокапсида в цитоплазму, а затем в ядро. Слияние происходит при низких значениях рН, которые создаются в эндоцитарной вакуоли — рецептосоме. Транскрипция и репликация генома происходят в ядре в составе нуклеокапсида.
Затравкой для транскрипции является КЭП, «шапочка», которая вместе с 10—13 нуклеотидами отрезается от клеточных и РНК, находящихся в ядре (отсюда необходимость ядерной локализации транскрипции), и перебрасывается на 5'-конец вирусного транскрипта.
На ранней стадии с наибольшей скоростью синтезируются иРНК для белков NS и NP, на поздней стадии — иРНК для белков НА, NA, М. Соответственно белки NS и NP являются ранними, а НА, NA и М — поздними. Белки Р синтезируются на протяжении инфекционного цикла с одинаковой скоростью.
Нуклеокапсиды транспортируются из ядра в цитоплазму и затем к клеточной оболочке к тем ее модифицированным участкам, на которых снаружи находятся вирусные гликопротеиды, а со стороны цитоплазмы — Mi-белок. Вирус выходит из клетки путем почкования.
Патогенез и клиника. Вирус гриппа попадает в организм через дыхательные пути вместе с каплями влаги и частицами пыли. Чем меньше величина капель и частиц, тем глубже проникает вирус в дыхательные пути. Благодаря короткому инфекционному циклу вируса (6—8 ч) при попадании в верхние дыхательные пути одной вирусной частицы уже через 8 ч количество инфекционного потомства достигает 103, а к концу первых суток — 1027. Эти расчеты объясняют столь короткий инкубационный период при гриппе — 1—2 сут. Репродукция вируса происходит в клетках эпителия слизистой оболочки дыхательных путей.
Может развиться первичная пневмония с некрозом Эпителия бронхиол. Пораженные клетки отторгаются, а продукты их распада всасываются, попадают в кровь, вызывая интоксикацию организма и лихорадочное состоя
ние. Вирус проникает в кровь и разносится по всему организму. Активация вирусом всей системы протеолиза и повреждение клеток эндотелия капилляров приводит к повышенной проницаемости сосудов, кровоизлияниям и дополнительному повреждению ткани. Возникают патологические изменения в почках, мозге. В результате повышенной проницаемости сосудов может возникнуть отек мозга с летальным исходом. Это случай так называемого молниеносного гриппа, когда смерть наступает на 2—3 сут. Вирус гриппа, попадая в кровь, вызывает угнетение кроветворения и иммунной системы, развивается лейкопения, может присоединиться интеркурентное заболевание, наблюдаются осложнения, вызванные бактериями и другими вирусами — затяжные риниты, гнойные синуситы, отиты, вторичные вирусные бронхиты и. пневмонии.
Однако описанное беспрепятственное распространение вируса в организме имеет место далеко не во всех случаях и может быть приостановлено при включении в процесс факторов неспецифического и специфического иммунитета, таких как неспецифические ингибиторы, находящиеся в крови, имеющиеся у больных антитела, интерферон. В результате развитие вирусной инфекции замедляется или прерывается, а через несколько суток появляются специфические антитела класса IgM, которые «защищают» организм. Антитела класса IgG появляются лишь спустя 2 нед после начала заболевания и их роль состоит в защите против повторного заражения данным серовариантом вируса гриппа.
Для клинического течения гриппа характерно быстрое развитие инфекции с высокой температурой, общей интоксикацией, воспалительными процессами в дыхательных путях. Температура достигает 38—39° С и более. Общая интоксикация выражается в головной боли, боли в глазных яблоках, резком угнетении. Развиваются симптомы местного поражения дыхательных путей — насморк, кашель, боли за грудиной. Поражение эндотелия стенок кровеносных сосудов приводит к точечным кровоизлияниям в верхних дыхательных путях, трахее, бронхах. Если развивается первичная гриппозная пневмония, она имеет геморрагический характер, и мокрота содержит прожилки крови.
Иммунитет. Пассивный иммунитет, передаваемый от матери, сохраняется у детей на протяжении первых месяцев жизни. После перенесенного заболевания возникает стойкий иммунитет.
Повторные заболевания гриппом обусловлены не кратковременностью иммунитета, а появлением новых серовариантов вируса гриппа, против которых у населения нет иммунитета.
В иммунитете против гриппа большую роль играют антитела класса IgA, накапливающиеся в секрете слизистой оболочки полости носа.
Эпидемиология. Эпидемиология гриппа своеобразна и не имеет аналогов среди других инфекционных заболеваний. Грипп передается воздушно-капельным путем. Больной или носитель (часть зараженных переболевает гриппом в инаппарантной или стертой форме) выделяют вирус вместе со слизью носоглотки и верхних дыхательных путей не только при кашле и чиханье, но и при разговоре. Так как инкубационный период болезни очень короток и не превышает 1—2 дней, то возникшая эпидемия распространяется довольно быстро, охватывая все новые и новые восприимчивые контингенты населения. Дальнейшее развитие эпидемии регулируется иммунитетом, который развивается после болезни через 1—1 '/2 нед, и поэтому по мере нарастания иммунной прослойки эпидемия гриппа идет на убыль. В условиях даже большого города на это уходит около месяца, а при нынешних широких связях между странами и быстрых транспортных средствах эпидемии гриппа в течение немногих месяцев охватывают население многих стран, нередко — все полушарие или даже всю планету. Такие обширные эпидемии называются пандемиями.
Причиной повторных эпидемий гриппа и повторных заболеваний является уникальная изменчивость белков вирусов гриппа (гемагтлютинина и нейраминидазы) в результате шифта и дрейфа. Люди, болевшие ранее гриппом, полностью восприимчивы к новому сероварианту вируса, к которому нет коллективного иммунитета, он начинает безудержно распространяться среди населения всего земного шара, вызывая пандемию гриппа.
' Первая достоверно документированная пандемия гриппа (ретроспективно мы знаем, что это был грипп типа А) возникла в 1889 г. Предполагается, что она началась в Китае, а затем в течение ближайших 1 '/2— 2 лет распространилась на все страны мира.
Следующая пандемия гриппа возникла через 29 лет — в 1918 г. За два года она распространилась во все страны мира и от нее погибло около 20 млн. человек. Никогда до этого и после этого грипп не протекал так тяжело.
На основании иммунологических исследований было установлено, что пандемия была вызвана вирусом, циркуляция которого к началу 30-х годов прекратилась среди людей и вирус был выделен от свиней. Эта пандемия началась в Китае, но он получил название «испанского» гриппа, так как первые упоминания о тяжелой эпидемии появились в Испании. Вирус, вызвавший эту эпидемию, в настоящее время обозначается H1N1. После 1918 г. эпидемии гриппа типа А повторялись через 2—3 года, а гриппа типа В — через 4—6 лет. В 1947—1949 годах возникла пандемия гриппа типаА, которая медленно распространялась и в течение 3 лет прошла во всех странах мира. Следующая пандемия получила название азиатской, а вызвавший ее вирус назвали вирусом азиатского гриппа. Эта пандемия, как и две предшествовавшие, началась в Китае (первый вирус, однако, был выделен в Сингапуре) в 1957 г. и к концу года охватила весь мир, вызвав заболевание от 1,5 до 2 млрд. человек. Пандемия была вызвана новым серовариантом вируса гриппа с антигенной формулой H2N2 (вирус азиатского гриппа). Одновременно перестал циркулировать вирус H1N1.
В дальнейшем эпидемии гриппа стали наблюдаться почти ежегодно, отличаясь степенью интенсивности — в зимние месяцы в северном полушарии, в летние месяцы — в южном полушарии. Новая пандемия разразилась через 11 лет, в 1968 г. она также началась в Китае и была вызвана вирусом, получившим название «гонконгского» по месту его выделения. Пандемия развивалась стремительно и поразила не менее 1 млрд. человек. Пандемию вызвал новый серовариант — гонконгский вирус гриппа с антигенной формулой H3N2, и одновременно перестал циркулировать среди людей азиатский вирус гриппа H2N2.
В 1977 г. произошло необычайное событие — «возвратился» вирус H1N1 после 20-летнего отсутствия. И на этот раз эпидемия началась в Китае. Заболевали почти исключительно лица моложе 20 лет, т. е. родившиеся после 1957 г., когда прекратилась циркуляция этого вируса среди населения. Другой особенностью явилось то, что предшествующий вирус (H3N2) не исчез, а также продолжал циркулировать и были выделены штаммы-рекомбинанты, содержавшие гены обоих вирусов. Вирус, вызвавший пандемию, получил название А/СССР/77, так как он впервые был выделен советскими учеными и немедленно передан ВОЗ. Позже выяснилось, что сходные
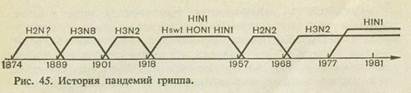
вирусы были выделены летом 1977 г. в Китае. История пандемий показана на рис. 45.
Начав циркулировать среди населения, новый вариант вируса не остается стабильным: его гликопротеиды НА и NA из года в год претерпевают антигенный дрейф, который приводит к тому, что через 2—3 года, а иногда и ранее выработанный иммунитет обеспечивает лишь частичную защиту от заболевания. Коллективный иммунитет (иммунологический пресс) является тем фактором, в результате которого селекционируются новые антигенные варианты, т. е. является движущей силой антигенного дрейфа.
По основному вопросу — источнику появления новых вариантов вируса гриппа в результате шифта — существуют две основные концепции. Согласно одной из них варианты, исчезнувшие из человеческой популяции, на самом деле продолжают циркулировать среди населения. Фактов, подтверждающих это предположение, нет. Другая гипотеза объясняет появление новых антигенных вариантов возвращением в человеческую популяцию вирусов, циркулирующих у животных. Во время эпидемии человек массивно выделяет вирус гриппа в биосферу и эпидемии и пандемии гриппа обычно сопровождаются эпизоотией среди домашних и диких животных, особенно птиц. Установлены экологические и эволюционные связи между вирусами человека и животных, и (благодаря секвени-рованию гена гемагглютинина) природа шифтов, которые привели к появлению азиатского и гонконгского серо-вариантов вирусов. Азиатский вирус сохранил четыре гена своего предшественника H1N1 — гены р|, Р2, Рз и NP, и получил новые четыре гена от предполагаемого партнера — гены НА, NA, M и NS. У гонконгского вируса H3N2 остались неизменными семь генов предшественника и только один ген НА был заменен в ходе рекомбинации. Возможными источниками этого гена были названы два вируса: вирус Hav7Neq2, выделенный от уток на Украине, и вирус Heq2Neq2, выделенный от лошадей в Майами (США).
Лабораторная диагностика. Материалом для исследования является отделяемое носоглотки, которое берут в первые 3 дня болезни ватными или марлевыми тампонами с задней стенки глотки, и мазки-отпечатки слизистой оболочки носовой полости. В летальных случаях используют кусочки пораженной легочной ткани, соскоб слизистой оболочки трахеи и бронхов.
Быстрая диагностика основана на выявлении вирусного антигена в эпителии слизистой оболочки верхних дыхательных путей. Наиболее широко используют метод ИФ. Антиген выявляют в мазках-отпечатках или эпителии, полученных из отделяемого носоглотки с помощью прямого и непрямого методов, используя коммерческие флюоресцирующие иммуноглобулины. Специфический антиген обнаруживается в цитоплазме в виде ярко светящихся конгломератов. Серологический вариант вируса гриппа (типов А, В или С) можно определить с помощью ИФ. Разработаны методы быстрой диагностики с помощью непрямой иммуноферментной реакции, основанные на идентификации типоспецифических антигенов (M и NP) в смывах носоглотки больных.
Выделение вируса. В связи со сменой серо-вариантов вируса гриппа желательно выделение его при вспышках и эпидемиях. Основным методом выделения вируса гриппа из смывов носоглотки является заражение 10—11-дневных куриных эмбрионов в амнион.
Идентификацию вируса проводят в РТГА или в РСК.
В сыворотках животных содержится большое количество неспецифических ингибиторов гемагглютинации, как термолабильных, которые разрушаются при прогревании сывороток при температуре 58° С, так и термостабильных. Вирусы гриппа могут быть как чувствительными, так и устойчивыми к ингибиторам. В первом случае иммунные сыворотки для удаления ингибиторов обрабатывают углекислотой, перийодатом калия или фильтратом холерного вибриона. Вирусы гриппа агглютинируют эритроциты с разной интенсивностью (авидностью) и в зависимости от этого их делят на авидные и неавидные штаммы. Для неавидных штаммов лучше использовать эритроциты человека группы 0. Для выделения вируса используют также культуры клеток эмбриона человека (почек и легких), почек обезьяны, МДСК и др. Индентификацию вируса проводят в РТГА, ИФ.
Серологическая диагностика заключается в обнаружении прироста антител в парных сыворотках больных. Используют РСК и РТГА, в качестве антигенов используют стандартные диагностикумы. Во второй сыворотке больного может быть обнаружен прирост антител сразу к двум или большему количеству используемых антигенов. В этом случае возбудителем заболевания считают тот штамм, к которому во второй сыворотке выявлен наивысший титр антител. Серологическую диагностику осуществляют также с помощью РН и ИФ в культуре клеток, реакции преципитации в геле, РРГ, ИФА. В РН используют наиболее чувствительные к вирусу культуры клеток почек собаки (MDCK) или быка (MDBK), а также клетки почек человека и обезьяны.
Профилактика и лечение. Применяются как убитые, так и живые вакцины. Те и другие имеют свои преиму-.щества и недостатки. Получение вакцинных штаммов основано на следующих принципах. Заранее приготавливается вирус-носитель, который обладает всеми необходимыми свойствами и хорошо размножается в лабораторных условиях (куриных эмбрионах). Используя технику рекомбинаци, от «актуального» вируса вирусу-носителю пересаживают только два гена — НА и NA, определяющих иммунологические свойства вируса. Для получения убитой вакцины вирус выращивают обычно на куриных"' эмбрионах, затем очищают от примесей, концентрируют и инактивируют, формальдегидом или другими химическими и физическими воздействиями, например, ультрафиолетовым облучением. При правильном подборе вакцинных штаммов вакцины снижают заболеваемость среди привитых в 2'/2—3 раза по сравнению с непривитыми. В качестве вируса-носителя для живых вакцин перспективно использование «холодовых» мутантов, которые неспособны к репродукции при температуре 37° С, но хорошо размножаются при более низких температурах, свойственных слизистой оболочке носоглотки (32—34° С). При этом создаются условия для репродукции вакцинного штамма в эпителии носоглотки, а не легочных альвеол. Для детей младшего возраста, а также лиц пожилого возраста, страдающих хроническими заболеваниями, разрабатываются вакцины со сниженной реактогенностью. Такие вакцины получают путем расщепления вирионов и выделения гемагтлютинина и нейра-минидазы (субъединичные вакцины). Для лечения и профилактики гриппа применяют химиопрепараты — аман-тадин и ремантадин. Последний является менее токсичным препаратом, его назначают внутрь в дозе 150—300 мг
в сутки в течение 2—3 дней. Лечение следует начинать как можно раньше после начала заболевания. Для лечения и профилактики гриппа используют интраназальное введение интерферона.
|
|
|
|
|
Дата добавления: 2014-11-20; Просмотров: 1137; Нарушение авторских прав?; Мы поможем в написании вашей работы!