КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Механизмы превращения протоонкогена в онкоген
|
|
|
|
Превращение протоонкогена в активно действующий онкоген обеспечивается следующими механизмами.
1. Присоединение к протоонокгену промотора – участка ДНК, с которым связывается РНК-полимераза, инициирующая транскрипцию гена, в том числе и онкогена, располагающегося непосредственно за ним (рис.1). Такого рода участки (промоторы) содержатся в больших терминальных повторах (LTR) ДНК-копий РНК-содержащих вирусов. Роль промотора могут выполнять и транспозирующие элементы генома – мобильные генетические элементы, способные перемещаться по геному и встраиваться в различные его участки.
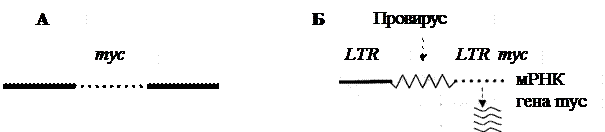 |
Рис. 1. Схема процесса активации протоонкогена в результате вставки промотора (по R.K. Marray et al.):
неактивный в нормальной хромосоме кур ген туc (А), оказавшись рядом (Б), вслед за LTR провируса лейкоза птиц после его интеграции в геном клетки активируется
2. Вставка в геном клетки энхансера (enchancer – усилитель) – участка ДНК, способного активизировать работу структурного гена, находящегося не только в непосредственной близости от него, но и на расстоянии многих тысяч пар нуклеотидов или даже встроенного в хромосому после него. Свойствами усилителя обладают подвижные гены, LTR ДНК-копий. В случае, проиллюстрированном рис. 2, LTR провируса не может работать как промотор (см. рис. 1, Б) и выступает в роли энхансера, в результате чего ген тус активизируется и транскрибируется.
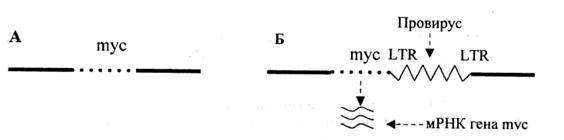
Рис. 2. Схема процесса активизации протоонкогена в результате вставки энхансера (по R.K. Marray et al., 1988):
А –нормальная хромосома кур, содержащая неактивный ген,mуc; Б –встраивание вируса лейкоза птиц в хромосому (в форме провируса) за myc-геном; активация и транскрибция гена туc.
|
|
|
3. Хромосомные абберации с явлениями транслокации, роль которых в механизмах опухолевой трансформации клетки можно проиллюстрировать следующим примером.
При лимфоме Беркитта конец q-плеча хромосомы 8, отделившись от нее, переходит к хромосоме 14: гомологичный фрагмент последней перемещается к хромосоме 8; а неактивный ген туc (протоонкоген), находившийся в том ее сегменте, который попадает на хромосому 14, встраивается вслед за активными генами, кодирующими тяжелые цепи молекул иммуноглобулинов, и активизируется (рис. 3, 4). Явления реципрокной транслокации между 9-й и 22-й хромосомами имеют место в 95 % случаев миелоцитарного лейкоза. Хромосома 22 с укороченным в результате такой транслокации одним плечом получила название Филадельфийской.
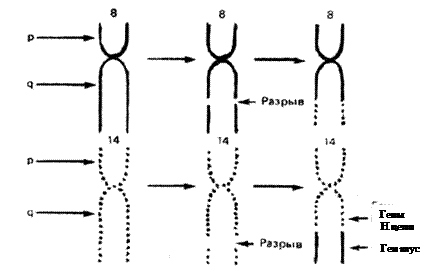 |
Рис. 3. Схема реципрокной транслокации в клетках лимфомы Беркитта (пояснение в тексте)
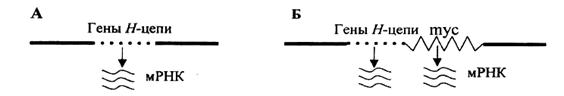
Рис. 4. Схема процесса активизации mус-протоонкогена при транслокации в клетках лимфомы Беркитта (по R.K. Муггау, 1988):
А – сегмент хромосомы 14 перед транслокацией, содержащий гены, кодирующие синтез иммуноглобулинов; Б – первично неактивный ген туc после транслокации оказывается под контролем энхансера, локализующегося в области генов, которые кодируют тяжелые цепи иммуноглобулина. В результате ген туc активизируется.
4. Точечные мутации протоонкогена, к примеру, C-H-raS, согласно
имеющимся сведениям, отличается от нормального гена (C-H-raS) всего одной
аминокислотой, но тем не менее обусловливает снижение гуанозинтрифосфатазной активности в клетке, что может вызвать рак мочевого пузыря у человека.
5. Амплификация (умножение) протоонкогенов, обладающих в норме
небольшой следовой активностью, обусловливает увеличение их общей активности до уровня, достаточного для инициации опухолевой трансформации. Известно, что в икринке шпорцевой лягушки около 5 млн копий гена туc. После оплодотворения и дальнейшего деления яйцеклетки число их прогрессирующе уменьшается. В каждой клетке будущего головастика в эмбриональный период развития содержится не более 20-50 копий myc-гена, обеспечивающих быстрое деление клеток и рост эмбриона. В клетках же взрослой лягушки выявляются лишь единичные гены туc, в то время как в раковых клетках той же лягушки число их вновь достигает 20-50.
|
|
|
6. Трансдукция неактивных клеточных генов (протоонкогенов) в геном ретровируса и последующее их возвращение в клетку: считается, что онкоген опухолеродного вируса клеточного происхождения; при инфицировании животных или человека таким вирусом «похищенный» им ген попадает в иной участок генома, что и обеспечивает активизацию некогда «молчавшего» гена.
|
|
|
|
|
Дата добавления: 2014-12-27; Просмотров: 2478; Нарушение авторских прав?; Мы поможем в написании вашей работы!