КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Потребность в восприятии и исследовании
|
|
|
|
10 12
3,0
К f,G
О
■О
1 «
6 8 Дна
Рис. 2. Кривая научения (отношение неправильных ответов к правильным) для 5 элементов механической головоломки за 12 дней эксперимента Головоломка состояла фактически из 5 элементов, шестой — шарнирная петля — был несъемным. Результаты 1-го, 2-го и 4-го дней значимо отличаются от результатов 10-го, 11-го и 12-го дней, р = 0,01. В конце каждого 2-часового сеанса экспериментатор входил в клетку и фиксировал число разобранных к этому моменту механизмов. Величина активности обезьян в ненаблюдаемый период определялась путем вычитания из этого числа количества механических элементов, разобранных в течение первых 5 мин. Эта величина также возрастала в течение 12 дней1
снова собрать головоломку, которую обезьяна успела разобрать в перерыве между его посещениями. В течение первых 5 мин каждого периода экспериментатор наблюдал за поведением обезьяны. Он отмечал, в частности, число правильных и неправильных манипуляций с игрушкой, проделанных обезьяной за это время. Действия считались правильными, если они приводили к разбору или освобождению одного из элементов головоломки, и неправильными, если обезьяна манипулировала с элементами не в установленном порядке или не могла их освободить.
На рис. 2 представлена кривая успехов двух обезьян, принимавших участие в опытах в течение 12 дней подряд; по ординате откладывается отношение неправильных действий к правильным. Кривая показывает, что научение обезьян происходит, несмотря на отсутствие какого-либо вознаграждения (пищи, например) извне. Другие графики автора свидетельствуют о том, что эти две обезьяны осуществляли соответственно 63 и 82 правильных действия в течение первых шести дней и 157 и 134 в течение последующих шести дней (за первые пять минут каждого сеанса). Интенсивность мотивации и активности обезьяны проявлялась, в частности, в том, что в последние б дней 81% головоломок разбирались практически в первые 5 мин после предъявления их животному.
|
|
|
На 13-й день Харлоу изучал феномен насыщения — каждые 6 мин в течение 10 ч (всего 100 сеансов) он предлагал каждой из обезьян вновь собранную головоломку. Он отметил заметное сокращение числа манипуляций, что свидетельствует об эффекте насыщения. Однако каждая из двух обезьян разобрала почти половину из возможного числа механизмов
 1 См.: Harlow H.F. Learning and satiation of response in intrinsically motivated complex puzzle performance by monkeys // J. Сотр. Physiol. Psychol. 1950. 43. P. 292.
1 См.: Harlow H.F. Learning and satiation of response in intrinsically motivated complex puzzle performance by monkeys // J. Сотр. Physiol. Psychol. 1950. 43. P. 292.
Нюттен Ж. [Познавательная и социальная мотивации] 117
 (100X6), а именно 286, что говорит об удивительной активности обезьяны и предполагает сохранение связанных с ней мотивов.
(100X6), а именно 286, что говорит об удивительной активности обезьяны и предполагает сохранение связанных с ней мотивов.
Б) Первичная потребность или производная тенденция? Пытаясь найти объяснение этой очень сильной мотивации, авторы предположили в своей первой работе1, что она имеет социальный характер из-за присутствия экспериментатора в начале каждого сеанса. Однако в конечном счете они пришли к отрицательному выводу и даже обнаружили, что у двух из четырех участвовавших в эксперименте обезьян уход экспериментатора в большей степени стимулировал активность животных, чем его присутствие. Возможность вторичного подкрепления, связанного с потребностью в пище, также исключается. Таким образом, авторам пришлось признать существование особой потребности в манипулировании. Харлоу2 обнаружил также, что удаление некоторых участков головного мозга приводит к снижению активности, связанной с манипулированием. Некоторые исследователи из школы Халла тем не менее подвергают сомнению специфический и первичный характер потребности в исследовании и манипулировании3. Следует отметить также, что до сих пор ни один из ученых не изучил происхождения и развития этой потребности у животных. С другой стороны, до сих пор точно не известны те свойства, которые придают новому стимулу положительную валентность для человека и животного. О более поздних исследованиях в этой области сообщается в работах Биндры и Спиннера, Карра и Брауна, Эрлиха4.
|
|
|
В) Амбивалентность необычного и неизвестного. При некоторых условиях неизвестное, новое или необычное вызывает негативную реакцию, причем ярко выраженные реакции страха наблюдаются значительно чаще, чем реакции приближения. В отдельных случаях имеет место любопытное сочетание притягательности и страха — тот соблазн опасного, но увлекательного приключения, который столь характерен для некоторых детских игр и поступков взрослых, стремящихся исследовать не-известное. Виз5 изучал негативные реакции ребенка при встрече с чужими или странными людьми. Он проследил развитие таких реакций от полного двигательного торможения и перевозбуждения до постепенного
 1 См.: Harlow H.F., Harlow M.K.,Meyer D.R. Learning motivated by a manipulation
1 См.: Harlow H.F., Harlow M.K.,Meyer D.R. Learning motivated by a manipulation
drive // J. Exp. Psychol. 1950. 40. P. 232-233.
2 См.: Harlow H,F. Learning and satiation of response in intrinsically motivated complex
puzzle performance by monkeys // J. Сотр. Physiol. Psychol. 1950. 43. P. 289-294.
3 См., например: Brown J.S. The motivation of behavior. N. Y.: McGraw-Hill Book,
1961. P.331-339.
4 См.: Bindra D., Spinner N. Response to different degrees of novelty: the incidence of
various activities // J. Exp. Anal. Behav. 1958. 1. P. 341-350; Carr R.M., Brown W.L. The
effect of sustained novelty upon manipulation on rhesus monkeys // J. Genet. Psychol.
1959. 61. P. 121-125; Ehrlich A. Effect of past experience on exploratory behavior in rats
// Canad. J. Psychol. 1959. 13. P. 248-254.
5 См.: Lewin K. A dynamic theory of personality. Selected papers. N. Y: McGraw-Hill,
1935. P. 261-264.
Тема 12. Психология мотивации
 привыкания в зависимости от расстояния до незнакомого человека и времени его присутствия и от того, находится ли ребенок в поле зрения или позади этого человека. С другой стороны, Хебб и Ризен1 изучали реакции страха у обезьяны, оказавшейся в незнакомой или новой обстановке. Выросший в изоляции шимпанзе не испытывает страха при первой встрече с человеком, хотя человеческое лицо, несомненно, является для животного «необычным» объектом. Но у той же самой обезьяны обнаруживаются реакции страха при виде нового человека после того, как она познакомилась с рядом людей и научилась их узнавать. Это нельзя объяснить ни образованием условных связей, ни генерализацией опасного стимула, поскольку до сих пор человек был для молодой обезьяны источником лишь приятных ощущений. Страх возникал у шимпанзе даже в том случае, когда кто-либо из известных уже ей людей приближался к клетке в одежде другого, также знакомого ей человека. На основании этих фактов Хебб2 сформулировал следующую гипотезу: расхождение между тем, что воспринимают рецепторы в данный момент, и следами того, что было воспринято ранее, приводит к определенному нарушению мозговых процессов, в результате возникает эмоция страха. Некоторые эксперименты3 поставили под сомнение универсальность этой гипотезы. Все это говорит о том, что проблема валентности неизвестного и непривычного по-прежнему остается нерешенной.
привыкания в зависимости от расстояния до незнакомого человека и времени его присутствия и от того, находится ли ребенок в поле зрения или позади этого человека. С другой стороны, Хебб и Ризен1 изучали реакции страха у обезьяны, оказавшейся в незнакомой или новой обстановке. Выросший в изоляции шимпанзе не испытывает страха при первой встрече с человеком, хотя человеческое лицо, несомненно, является для животного «необычным» объектом. Но у той же самой обезьяны обнаруживаются реакции страха при виде нового человека после того, как она познакомилась с рядом людей и научилась их узнавать. Это нельзя объяснить ни образованием условных связей, ни генерализацией опасного стимула, поскольку до сих пор человек был для молодой обезьяны источником лишь приятных ощущений. Страх возникал у шимпанзе даже в том случае, когда кто-либо из известных уже ей людей приближался к клетке в одежде другого, также знакомого ей человека. На основании этих фактов Хебб2 сформулировал следующую гипотезу: расхождение между тем, что воспринимают рецепторы в данный момент, и следами того, что было воспринято ранее, приводит к определенному нарушению мозговых процессов, в результате возникает эмоция страха. Некоторые эксперименты3 поставили под сомнение универсальность этой гипотезы. Все это говорит о том, что проблема валентности неизвестного и непривычного по-прежнему остается нерешенной.
|
|
|
Стремление к визуальному восприятию и исследованию наилучшим образом было продемонстрировано в известных экспериментах Батлера4. Пять обезьян-резусов помещались в клетку с непрозрачными стенками, освещавшуюся изнутри лампочкой в 25 Вт. На передней стенке находились две небольшие дверцы (рис. 3), которые открывались посредством электрического механизма, как только обезьяна нажимала на одну из них. После предварительного периода тренировки перед дверцами вывешивались желтая и синяя картонки с тем, чтобы указать, ка-
1 См.: Hebb D.O., Riesen A.H. The genesis of irrational fears // Bull. Canad. Psychol.
Ass. 1943. 3. P. 49-50.
2 См.: Hebb D.O. The organization of behavior. A neuropsychological theory. N. Y.:
John Wiley & Sons, 1949. P. 149.
3 См.: Hunt J.McV., Quay H.C. Early vibratory experience and the question of innate
reinforcement value of vibration and other stimuli // Psychol. Rev. 1961. 68. P. 149-156;
Warren R.P., Pfaffmann С Early experience and taste aversion // J. Сотр. Physiol. Psychol.
1958- 52. P. 263-266; Meier G.W., Foshee D.P. et al. Helson's residual factor versus innate
R-S relations // Psychol. Rep. 1960. 6. P. 61-62.
4 См.: Butler RA. Discrimination learning by rhesus monkeys to visual-exploration
motivation // J. Сотр. Physiol. Psychol. 1953. 46. P. 95-98.
Июлей Ж, [Познавательная и социальная мотивации]
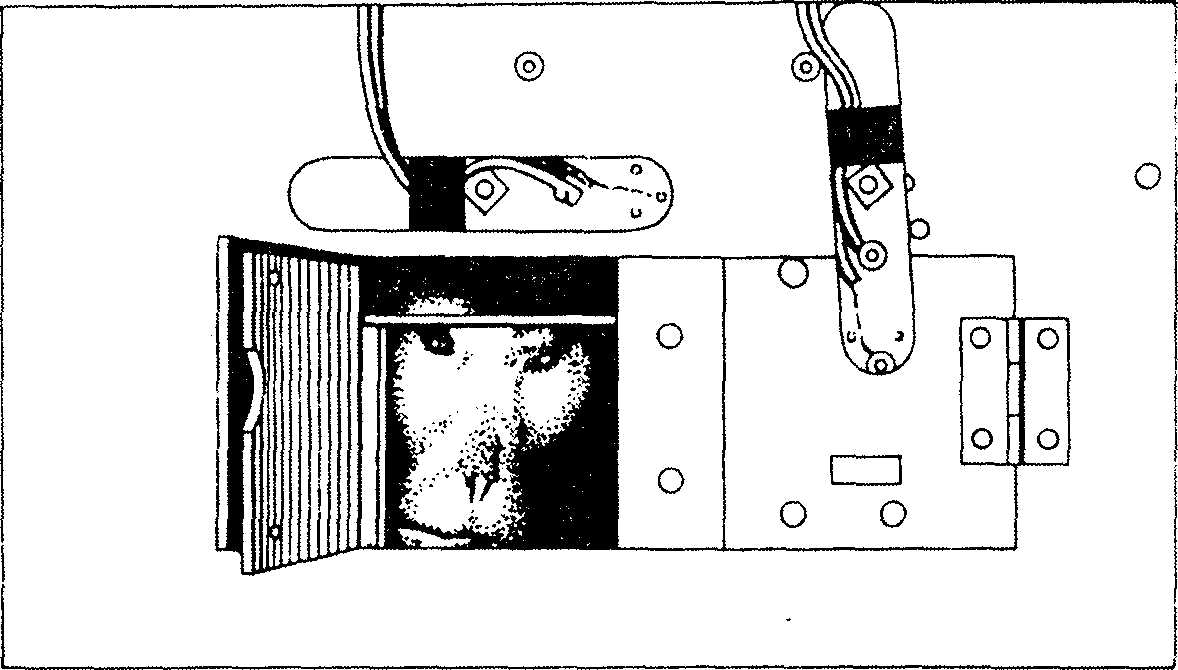
Рис. 3. Частичный вид устройства, применявшегося
для изучения визуальной исследовательской деятельности. Видны две дверцы, закрывающиеся и открывающиеся автоматически с помощью электрического устройства. Через открытую дверцу видна голова макаки-резус, исследующей окружающую обстановку1
|
|
|
кая из дверей автоматически откроется при нажатии на нее. Обезьяны, таким образом, обучались визуальному различению. Единственным вознаграждением обезьяне всякий раз, когда она нажимала на правильную дверцу, служило то, что, как только последняя открывалась, обезьяна получала возможность заглядывать через отверстие в лабораторию и наблюдать за тем, что там происходит. Через 30 с опускалась ставня. Если обезьяна нажимала на заблокированную дверцу, зажигалась маленькая лампочка и сразу же опускалась ставня, закрывавшая обе дверцы. За каждой пробой — как удачной, так и неудачной — следовал 30-секундный интервал, во время которого картонки менялись местами в случайном порядке, затем ставня поднималась, и обезьяна снова могла их видеть. Обезьяна располагала для выбора 5 мин. Если в течение этого времени она никак не реагировала на картонки, дверца открывалась извне, оставалась открытой в течение 30 с, затем опускалась ставня, и спустя 30 с снова предъявлялись картонки. Разумеется, пробы могли следовать с большей или меньшей частотой в зависимости от того, насколько быстро обезьяна нажимала на дверцу, после того как поднималась ставня. Каждая обезьяна делала 20 проб ежедневно на протяжении 20 дней (5 дней в неделю).
1 См: Butler ЯЛ., Harlow H.F. Persistence of visual exploration in monkeys // J. Сотр. Psysiol. Psychol. 1954. 47. P. 259.
Тема 12. Психология мотивации
| Обезьяна \Q2 | ||||||||
| 100 л | Ю0- | а^ибезьяна 147 | ||||||
| равильь | тов | 80- | < | правиль етов | 80~ 30- | ^и^С^лучайные результаты | ||
| hfj — | V | ^ "■* *"•■ *" "«^ •»» •■» "•» —- *-* | ||||||
| Е | 00 О | айные результаты | S i | 40- | ||||
| 40- | о | 20- | ||||||
| W | т, | * | ||||||
| 4 6 | 8 10 /2 И ffi 18 20 | Т!! 1 1 / \ 1 1 г 1 \ /23456789 1011 | ||||||
| Дни | Группы по 100 про5 |
Рис. 4а. Кривая научения 3 обезьян в опыте Батлера с визуальным различением, где мотивацией являлась
исследовательская деятельность1
Рис. 46. Индивидуальные кривые научения 2 других обезьян в опыте Батлера с визуальным различением, где мотивацией являлась исследовательская деятельность.
Показано увеличение числа правильных ответов на протяжении 11 последовательных серий по 100 проб в каждой. Обезьяна 102 достигла и удерживалась на уровне почти безошибочной деятельности в течение 600 последних проб. Результаты обезьяны 147 в последние 600 проб хотя и несколько хуже, однако значимо выше случайных (р = 0,01)2
Полученные результаты показывают, что все животные научились выполнять задачу на различение (рис. 4).
Исследователи измеряли также латентное время реакции (т.е. время от поднятия ставни до нажатия на одну из дверц). Были отмечены индивидуальные различия у 5 животных, участвовавших в этом эксперименте, но латентное время в последние дни опыта возросло (по сравнению с первыми днями) только в одном из пяти случаев. Следовательно, рассмотренная нами мотивация успешно противостояла насыщению3.
1 См.: Butler RA. Discrimination learning by rhesus monkeys to visual-exploration
motivation // J. Сотр. Physiol. Psychol. 1953. 46. P. 96.
2 См. там же.
3 См. так же: Butler RA., Harlow H.F. Persistence of visual exploration in monkeys //
J. Сотр. Psysiol. Psychol. 1954. 47. P. 258-263.
НюттенЖ. [Познавательная и социальная мотивации] 121
4. Теоретическая интерпретация
Харлоу1 подчеркнул различие между потребностью в исследовании и гомеостатическими потребностями. Он говорил о противоположности этих двух категорий потребностей: одни из них имеют внутреннее происхождение, другие — внешнее. Такое противопоставление кажется нам некоторым преувеличением, поскольку оно основывается на различном — внешнем или внутреннем — источнике мотивации. Действительно, наблюдения этологов показали важное значение внешних объектов (releasers, или пусковых механизмов) для развертывания поведения, что, однако, не умаляет значения органических или внутренних компонентов мотивационного фактора. С другой стороны, можно считать, что источником всякой мотивации является либо индивид, «имеющий определенную тенденцию», либо объект, обладающий валентностью.
Тем не менее нельзя, видимо, отрицать глубокого различия между гомеостатическими и вообще физиологическими потребностями и познавательными тенденциями (потребностью в исследовательской деятельности, восприятии, познании). Но искать это различие следует, скорее, в природе «завершающих реакций»2, характерных для того и другого типа потребностей и поведения. В то время как удовлетворение гомеостати-ческих потребностей связано, как правило, с действиями, оказывающими биологическое воздействие на организм или на его потомство, удовлетворение познавательных потребностей связано, видимо, с самим объектом. Отношения организма со средой предполагают, таким образом, некоторую активность, свидетельствующую о том, что организм на определенной ступени эволюции способен выделять некоторые объекты и интересоваться ими как таковыми. Видимо, именно здесь и следует искать источник могущественных объектных мотивов, или подчиненности объекту, столь характерных для некоторыхпоступков и помыслов человека, например для его научной деятельности. Келли3 справедливо подчеркивает, что такая установка, характерная для ученых, не является исключительно привилегией немногих избранных; каждый человек, говорит он, является одновременно man-the-scientist4, стремящимся познать мир таким, каков он есть, и man-the-bio-logical-organism5, на котором чрезмерно сосредоточен интерес психолога.
 1 См.: Harlow H.F. Mice, monkeys, men and motives // Psychol. Rev. 1953. 60. P. 23-32;
1 См.: Harlow H.F. Mice, monkeys, men and motives // Psychol. Rev. 1953. 60. P. 23-32;
Harlow H.F. Motivation as a factor in the acquisition of new responses // Current theory and
research in motivation. A symposium. Lincoln: Univ. of Nebraska Press, 1953. P. 24-49.
2 См.: Шеррингтон Ч. Интегративная деятельность нервной системы. Л., 1969.
С.311-314.
3 См.: Kelly СЛ. The psychology of personal constructs. N. Y.: W.W. Norton & Co.,
1955. Vol. 2. P. 4.
4 Человек-ученый {англ.).
5 Биологический организм {англ.).
122 Тема 12. Психология мотивации
5. Тенденция к чередованию
С потребностью в исследовании, с одной стороны, и феноменом насыщения — с другой, следует связать тенденцию к чередованию, отмеченную у животных, например, при выборе маршрута в лабиринте. Этот феномен можно описать следующим образом: животное, которое в Т-образном лабиринте поворачивает в первый раз направо, при второй пробе чаще всего поворачивает налево. Это происходит не только в опытах без подкрепления, но также и в том случае, если в конце первого коридора находится вознаграждение. Фаулер1 уточнил этот вопрос, указав, что количественно небольшое вознаграждение (пища) практически не влияет на тенденцию к чередованию, тогда как большое явно уменьшает ее. В последнем случае тенденция к повторному выбору первого пути берет верх над тенденцией к чередованию.
Этот феномен чередования был впервые отмечен Толменом2, а затем подвергнут систематическому изучению Деннисом3, Монтгомери и многими другими4. Это явление нельзя полностью объяснить реактивным торможением, как это пытался сделать Халл. Оно связано, скорее, как это показал, в частности, Монтгомери, с поиском новых стимулов и потребностью в исследовательской деятельности. Действительно, данный феномен является, видимо, упрощенной формой того, что наблюдается в более сложных ситуациях, например в лабиринте Дешиэлла5 с множественным выбором, когда животное выбирало маршруты, охватывавшие большую часть лабиринта. В лабиринте другого типа, в форме креста с двумя входами и двумя камерами на концах <...>, у животных отмечается отчетливая тенденция двигаться по очереди к каждой из камер, а не поворачивать попеременно то налево, то направо, как это требудет за-
1 См.: Fowler H., Fowler D.E., Dember W.N. The influence of reward on alternation behavior
// J. Сотр. Psysiol. Psychol. 1959. 52. P 220-224; Fowler H., Blond J., Dember W.N. Alternation
behavior and learning: the influence of reinforcement magnitude, number and contingency //
J. Сотр. Physiol. Psychol. 1959. 52. P. 609-614.
2 См.: Tolman E.C. Purpose and cognition: the determiners of animal learning // Psychol.
Rev. 1925. 32. P. 285-297.
3 См.: Dennis W. A comparison of the rat's first and second exploration of a maze unit.
// Amer J. Psychol. 1935. 47. P. 488-490; Montgomery K.C. The relation between exploratory
behavior and spontaneous alternation in the white rat // J. Сотр. Physiol. Psychol. 1951.
44. P. 582-589; Montgomery K.C. A test of two explanations of spontaneous alternation //
J. Сотр. Physiol. Psychol. 1952. 45. P. 287-293; Montgomery K.C. Exploraory behavior and
its relation to spontaneous alternation in a series of maze exposures // J. Сотр. Physiol.
Psychol. 1952. 45. P. 50-57.
4 См.: Dember W.N., Fowler H. Spontaneous alternation behavior // Psychol. Bull. 1958.
55. P. 412-428.
5 См.: Dashiell J.F. A quantitative demonstration of animal drive // J. Сотр. Psychol.
1925. 5. P. 205-208.
Нюттен Ж. [Познавательная и социальная мотивации] 123
 кон реактивного торможения1. Другими словами, животное не избегает повторения одного и того же движения (например, поворота налево), но оно старается не приходить два раза подряд в то же самое место и, как правило, идет туда, где оно еще не побывало.
кон реактивного торможения1. Другими словами, животное не избегает повторения одного и того же движения (например, поворота налево), но оно старается не приходить два раза подряд в то же самое место и, как правило, идет туда, где оно еще не побывало.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 490; Нарушение авторских прав?; Мы поможем в написании вашей работы!