КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Общая психопатология 17 страница
|
|
|
|
Достоверные частные коэффициенты корреляции получены при режиме «вдыхание СОг — гипервентиляция» (р = 0,286, р>> 0,998) и при режиме «гипервентиляция — уменьшение вентиляции» (f> = 0,357, р>>0,999). Это еще раз подтверждает зависимость изменений амплитуды пульсовых волн плетизмограмм в наших исследованиях от рСОг в альвеолах.
Для каждого из трех вышеуказанных режимов рассчитаны коэффициенты прямолинейной регрессии (В), показывающие, на сколько единиц изменяется амплитуда пульсовых волн плетиз-мограммы при изменении рСОгА на 1 мм рт.ст.
При гипервентиляции В = 1,1; при режиме «вдыхание С02 — гипервентиляция» В =1,47; при режиме «гипервентиляция — уменьшение вентиляции» В = 3,3.
Отрицательная зависимость между рСОгА и тонусом периферических артерий, полученная в наших исследованиях у больных гипертонической болезнью и коронарной недостаточностью, а также отрицательная корреляция между рСОгА и минимальным и средним динамическим АД, установленная при обработке исходных данных, дают возможность предполагать, что в патогенезе повышения сосудистого тонуса у больных коронарной недостаточностью и гипертонической болезнью может играть роль обнаруженное многими исследователями хроническое уменьшение парциального давления углекислоты в альвеолярном воздухе и артериальной крови.
Приведенные данные о влиянии углекислоты в альвеолярном воздухе на тонус артериальных сосудов у больных коронарной недостаточностью и гипертонической болезнью получены в результате кратковременного исследования. Патогенез гипертонической болезни сложен. Для получения более глубоких представлений о роли изменений парциального давления С02 в альвеолярном воздухе и артериальной крови нужны более длительные клинические наблюдения.
|
|
|
Выводы:
1. При уменьшении парциального давления углекислоты в альвеолярном воздухе вследствие гипервентиляции у больных коронарной недостаточностью и гипертонической болезнью происходит увеличение тонуса периферических артерий.
2. При плавном увеличении парциального давления углекислоты в альвеолярном воздухе вследствие вдыхания газовой смеси с повышенным содержанием СОг или в результате уменьшения легочной вентиляции у больных коронарной недостаточностью и гипертонической болезнью происходит уменьшение тонуса периферических артерий.
3. Вышеуказанные изменения тонуса сосудов обусловлены, по-видимому, главным образом соответствующими изменениями парциального давления углекислоты в альвеолярном воздухе.
4. Установлено наличие положительной корреляции между рСОг в альвеолярном воздухе и амплитудой пульсовых волн плетизмограмм при гипервентиляции, и при объединенных режимах (вдыхание СО2 — гипервентиляция, гипервентиляция — уменьшение вентиляции). Таким образом, получена количественная оценка зависимости тонуса периферических артерий от величины парциального давления углекислоты в альвеолярном воздухе.
5. При уменьшении парциального давления углекислоты в альвеолярном воздухе у части исследуемых обнаружены изменения ЭКГ, указывающие на увеличение гипоксии миокарда. При этом у больных наблюдаются отрицательные субъективные симптомы (головокружение, неприятные ощущения в области сердца).
6. В ответ на увеличение парциального давления углекислоты в альвеолярном воздухе вследствие вдыхания углекислоты или уменьшения легочной вентиляции у части исследуемых обнаружены изменения ЭКГ, свидетельствующие об уменьшении гипоксии миокарда. При этом полностью исчезают все отрицательные субъективные симптомы, вызванные уменьшением СОг в альвеолярном воздухе вследствие гипервентиляции.
|
|
|
7. По группе больных (п = 112) установлена отрицательная корреляция между парциальным давлением СОг в альвеолярном воздухе и минимальным, а также средним динамическим артериальными давлениями.
8. Определены средние «критические» уровни рСОг в альвеолярном воздухе, переход через которые в процессе исследования приводил к изменению тонуса мелких периферических артерий. При пробе с вдыханием CQL> средний «критический» уровень рСОг равен 42,1+0,16 мм рт.ст., при гипервентиляции — 31,7 + 0,1 мм рт. ст., при уменьшении легочной вентиляции — 38,5±0,1 мм рт. ст.
В. А. КАЗАРИНОВ
биохимическое обоснование теории болезни глубокого дыхания к. п. бутейко
ТЕОРЕТИЧЕСКИЕ ПРЕДПОСЫЛКИ МЕТОДА ВЛГД
I. РОЛЬ УГЛЕКИСЛОТЫ В ПРОЦЕССАХ ЖИЗНЕДЕЯТЕЛЬНОСТИ ОРГАНИЗМА (ХИМИЧЕСКИЕ И БИОХИМИЧЕСКИЕ АСПЕКТЫ)
1. Формы углекислоты
В результате многочисленных исследований стало известно, что углекислота в организме животных и человека является не только компонентом буферных систем и регулятором дыхания, но и фактором регуляции деятельности сердечной, сосудистой, гормональной, нервной и пищеварительной систем. В настоящее время можно сказать, что углекислота в организме животных и человека участвует в регуляции ряда важнейших физических функций.
Говоря об углекислоте, часто подразумевают только одну ее форму, а именно газообразную — СО2 (углекислый газ). При этом упускается из виду, что в организме углекислота находится в пяти различных формах:
| 2- |

|
1) растворенный углекислый газ (рССЬ);
2) угольная кислота (Н2СО3);
3) бикарбонаты (НСОГ);
4) карбонаты (СОз~);
2. Роль углекислоты в процессе дыхания
Основным источником углекислоты в организме является эндогенный углекислый газ. Причем в зависимости от различных факторов интенсивность образования С02 в тканях в процессах обмена может варьировать в значительных пределах. Однако концентрация СОг в крови и других биологических жидкостях организма в норме обычно не подвержена таким сильным изменениям. Как правило, она поддерживается на строго определенном уровне, а излишек немедленно удаляется. Последнее обеспечивается функционированием сложной системы механизмов регуляции ее концентрации. Важнейшим из них является процесс нейтрализации и быстрого выведения излишка С02 из тканей в окружающую среду. Этот механизм можно представить следующим образом.
|
|
|
Образующаяся внутри клетки СОг проникает в межклеточную жидкость, а затем в кровь. Механизм переноса С02 через клеточные мембраны не совсем ясен. Известно только, что мембраны более проницаемы для С02 и Н2С03, чем для НСОз~. По мере проникновения углекислоты в межклеточную жидкость, а оттуда в кровь, основная масса превращается в НСО.Г • Процесс катализируется карбоангидразой (КА) эритроцитов:
КА
С02 + Н20-------------- ^Н2СОз Н+ + НСО3-.
Вследствие этого процесса рН внутри эритроцита начинает понижаться. При определенных значениях рН имидозольная группа гистидина и аминогруппа валина цепей гемоглобина присоединяют Н +, в результате чего приобретают (+) заряд, что позволяет стабилизировать солевые мостики, которые отвечают за сохранность дезоксиструктуры гемоглобина. Помимо этого процесса идет процесс вытеснения СГ из сети электробатиче-ских взаимодействий углекислым газом, что приводит к повышению устойчивости четвертичной структуры гемоглобина. Кроме того, углекислый газ влияет на связывгкие ДФГ и присоединяется к а-аминогруппам, что также стабилизирует дезоксиструктуру гемоглобина (эффект Вериго — Бора).
За счет разности концентраций кислорода в эритроците и тканях кислород начинает диффундировать в ткани. Вследствие действия кооперативного эффекта связывания Ог отрыв каждых последующих молекул кислорода все более облегчается.
Суммарным результатом этих двух процессов будет поддержание рН практически постоянным, облегчение диффузии кислорода и удерживание внутри эритроцитов ионов К+, ранее нейтрализованных НЬОг, а теперь нейтрализуемых образовавшимися ионами НСО.Г.
В результате большая часть С02, которая диффундировала из тканей в эритроциты, переходит из капилляров в вены в виде НСО.Г, находящихся в эритроцитах.
Благодаря этому изокислотному сдвигу отношение НСО.Г эритроцита (э) к НСО.Г плазмы (п) изменяется за счет повышения НСО.Г в эритроцитах. Отношение НСО.Г э/С1~ э более не сохраняется равным отношению НСО.Г п/СГ п, которое устанавливалось за счет проявления эффекта Гиббса - Доннана. Тенденция НСО.Г покидать клетки увеличивается, а вышедшие ионы НСО.Г замещаются на CP из плазмы до тех пор, пока не будет достигнуто новое равновесие. Итоговым значением этого процесса является то, что значительная доля общего количества СО2, которая в крови капилляров попадает в эритроциты, теперь оказывается в виде НСО.Г в венозной плазме. Следует заметить, что хотя превращение НЬ02 в НЬН+ и последующее буферное действие этого превращения не влияют на осмотическое давление, обусловленное гемоглобином клеток, поскольку количество гемоглобина не меняется, суммарный эффект изо-кислотного сдвига и хлоридного сдвига приводит к увеличению общего количества анионов и благодаря этому к увеличению эффективного осмотического давления в клетках. Вследствие это-
|
|
|
го вода перераспределяется между клетками и плазмой.
В легких процесс идет в обратную сторону. За счет разности давлений кислород проникает внутрь эритроцита и образует кислоту ННЮ2. Вследствие ее диссоциации образуются ионы Н +, которые, взаимодействуя с НСО.Г, образуют Н2С03. Н2СОз под действием карбоангидразы расщепляется на С02 и Н20:
н2со3^н2о+со2.
С02 и Н20 выделяются в полость альвеол. Удаление С02 приводит к смещению реакции Бора в левую сторону, что опять-таки приводит к нормализации рН и облегчает связывание кислорода с гемоглобином.
Одновременно с этим процессом в легких идет процесс расщепления карбонатов и переход белков в свободное состояние.
Поступивший в кровь бикарбонат, взаимодействуя с угольной кислотой, образует пару НСОз~/Н2С03, которая является компонентом главного буфера внеклеточной жидкости. Необходимость существования именно такого буфера обусловлена многими причинами:
1) во внеклеточной жидкости присутствует значительно больше НСОГ, чем любого другого буферного соединения;
2) поступление С02 не ограничено;
3) физиологические механизмы, поддерживая нормальную величину рН внеклеточной жидкости, регулируют в ней концентрации либо НСО.Г, либо СОо. Van Slyke и затем Warburg нашли, что между рН, рС02 и НСОз" крови существует линейная зависимость, характеризуемая уравнением Гендерсона Гессельбаха:
 |
pH=6,l+lg
[НСО.1 1
[рСО,]
4) буферная система НСОз~/Н2С03 функционирует вместе с системой НЬ, как описано выше.
Как и у всех буферных систем, рН зависит не от абсолютных концентраций компонентов буфера, а от их отношения (табл. 1, 2, 3 из книги: Робинсон Дж. Р. Основы регуляции кислотно-щелочного равновесия. М., Медицина, 1969).
Таблица 1
Влияние рСОг на рН раствора, содержащего 27 м.-экв бикарбоната на I л при 37°С
| рСОг мм рт. ст. | 80 160 | |||
| Н2СО3 м.-экв/л | 0,3375 | 0,675 | 1,35 | 2,7 5,4 |
| НСОз- м.-экв/л | 27 27 | |||
| НСО3-/Н2СО3 | 10 5 | |||
| РН | 8,0 | 7,7 | 7,4 | 7,1 6,8 |
| Таблица 2 | ||||
| Влияние изменений концентрации бикарбоната | ||||
| рСОг мм рт. ст. | ■ | |||
| НгСОг м.-экв/л | 1,35 | 1,35 | 1,35 | |
| НСОГ м.-экв/л | 13,5 | |||
| НСО3-/Н2СО3 | ||||
| РН | 7,7 | 7,4 | 7,1 | |
| Таблица 3 | ||||
| Взаимодействие изменений рСОг | и НСОъ | |||
| рСОг мм рт. ст. | ||||
| Н2СО3" м.-экв/л | 2,7 | 0,765 | 0,3375 | |
| НСОз- м.-экв/л | 13,5 | 6,75 | ||
| НСО3-/Н2СО3 | ||||
| РН | - | 7,4 | 7,4 | 7,4 |
Однако следует сказать, что хотя в табл. 3 рН сохраняется неизменной, буферная емкость плазмы падает. Буферная эффективность системы НСОз значительно возрастает в присутствии эритроцитов.
Рассмотренное сочетание свойств буфера, одним из компонентов которого является газ, и автоматической саморегуляции, возможной за счет внутриклеточного Нв, обеспечивает замечательное постоянство рН плазмиды крови. Кроме того, организм использует еще две стабилизирующие системы -дыхательный аппарат и почки, которые, регулируя соответственно [Н2СО*] и [НСОз~], создают дополнительные возможности поддержания постоянства рН внеклеточной жидкости.
3. Регуляция уровней С02, НСОз и Н+ в тканях
животных
В отличие от [НСО.Г] (фиксируемой концентрации аниона) [НзСОл] определяется исключительно одним параметром: парциальным давлением С02 в газовой смеси, находящейся в равно-
весии с внеклеточной жидкостью, а именно в альвеолярном воздухе. Это давление в свою очередь зависит от скорости, с которой С02, выходящий из крови в легких, разбавляется атмосферным воздухом, а следовательно, от глубины и частоты дыхания. Характер дыхания регулируется дыхательным центром нервной системы; это центр, по-видимому, чувствителен к рН и рС02 внеклеточной жидкости. Когда рН внеклеточной жидкости падает ниже нормы из-за уменьшения [НСОг], дыхание стимулируется, что приводит к снижению альвеолярного рС02 и, следовательно, внеклеточной [Н2СОз] • Это ведет к возвращению соотношения [НСО.Г] / [Н2СОз] к нормальной величине 20:1 и к возвращению рН. Наступающее снижение напряжения С02 в плазме противоположным образом влияет на регулирующий нервный центр, следовательно, компенсация никогда не была бы полной, если бы функционировал только этот регуляторный механизм.
При высоком рН плазмы частота дыхания снижается, альвеолярное рС02 и, следовательно, [Н2СОз] в плазме возрастают, а рН сдвигается к норме. Полная компенсация при этом не достигается, так как повышенная [Н2СОз] в плазме действует на ды-
хательный центр противоположно влиянию на него увеличенного рН. Если частота дыхания значительно снижается, то уменьшение р02 будет стимулировать увеличение дыхательной активности.
В то время как дыхательный механизм компенсирует нарушения кислотно-щелочного равновесия путем регуляции Н2С03] во внеклеточной жидкости, почки участвуют в контроле рН путем регуляции [НСОз-]. Снижению внеклеточного рН, обусловленного увеличением альвеолярного рС02 или уменьшением [НСО.Г], почки противодействуют двумя доступными им путями, а именно выделением Н+ в форме либо недиссоциирован-ной кислоты, либо NH^.
Почечная компенсация при обстоятельствах, которые могли бы привести к повышению внеклеточного рН, осуществляется путем снижения [НСО.Г] внеклеточной жидкости.
Имеются данные о том, что одновременно с этими механизмами, компенсирующими колебания рН во внеклеточной жидкости, срабатывает и «калий-натриевый насос». Мышечные клетки, клетки эпителия почечных канальцев, а возможно, и все другие клетки обладают ионообменным механизмом, который осу-
ществляет обмен через клеточную мембрану либо К, либо Н, либо обоих этих ионов на Na +. Благодаря этому обмену содержимое клетки может участвовать в поддержании внеклеточного рН.
При увеличении внеклеточной [НСОз-], Na + поступает в клетки в обмен на Н-1" и К +. Протоны реагируют с внеклеточными НСОз". и образующийся С02 удаляется с выдыхаемым воздухом, К4" выделяется с мочей вместе с эквивалентным количеством НСОз". При уменьшении рН Na4" выходит из клетки, а ионы Н+ К+ зходят в нее.
По направлению сдвига активной реакции крови все изменения кислотно-щелочного баланса делятся на ацидоз и алкалоз, а по причине, вызывающей изменения рН крови, на метаболические и респираторные.
4. С д в и г и кислотно-щелочного равновесия
Ацидоз респираторный. Характеризуется следующими показателями: рСО>>нормы; рН<7,36; [НСОз-] норма или больше нормы. Эти состояния наблюдаются при подавлении деятельности дыхательного центра морфием, при вдыхании газовой смеси, содержащей большой процент углекислоты, при тяжелых формах пневмоний, отеке легких и эмфиземе, при самопроизвольной задержке дыхания, астме, при удушье.
Алкалоз респираторный. Показатели: рСОг<нормы; pH>7.44; |НСО; —норма или несколько меньше нормы. Развивается это состояние при гипервентиляции легких, при острых воспалениях внутренних половых органов, гипоксиях и недостаточности кровообращения. При длительно существующем респираторном алкалозе постоянно наблюдается увеличение молочной кислоты, которая образуется вследствие недоокисления пи-рувата, т. е. в отсутствие достаточных концентраций кислорода в тканях.
Апидоз метаболический. Показатели: рН<нормы; [НСО.Г ]< нормы; рС02<нормы. Развивается в случаях появления избыточного количества нелетучих кислот во внеклеточной жидкости, которое появляется:
1) вследствие приема пищи с большим количеством нелетучих кислот;
2) вследствие образования большого количества органических кислот (молочная, ацетоуксусная, пировиноградная и др.) при недостатке кислорода в тканях;
3) вследствие нарушения функций почек по выведению ионов водорода;
4) при относительном избытке нелетучих кислот, вследствие потери оснований при рвоте, поносе, желчных свищах и т. д.
Алкалоз метаболический. Показатели: повышение рН; [НСО.Г] > нормы; рСО_>> нормы. Развивается в случае потери нелетучих кислот или в случае потери К+ (что приводит к избыточному выведению Н+ почками).
Широкое распространение указанных состояний организма животных и человека стало главной причиной того, что еще с конца XIX века и по настоящее время многочисленные исследования посвящаются изучению особенностей обмена веществ в тканях при различного рода алкалозах и ацидозах. Однако подходы к изучению этой проблемы и объяснение полученных результатов длительное время основывались в большинстве случаев только на роли изменения концентрации ионов водорода и почти совершенно не обращалось внимания на важное метаболическое значение изменения содержания в тканях различных форм углекислоты при этих состояниях.
За последние годы интерес к выяснению метаболического значения углекислоты возрос. Было установлено, что она принимает участие в регуляции многочисленных узловых звеньев обмена веществ и соответственно физиологических функций организма. В частности, было показано, что интенсивность процессов биосинтеза основных органических компонентов ткани
животных находятся в прямой зависимости от интенсивности процессов фиксации углекислоты (реакции карбоксилирования).
5. Реакции карбоксилирования
Еще до недавнего времени не было известно, что процессы фиксации углекислоты лежат в основе всех важнейших обменных и, в первую очередь, биосинтетических процессов в тканях. В настоящее время по этим вопросам опубликован ряд статей. Суммируя накопленные данные, можно заключить, что в тканях организма существует большое количество реакций, которые в
зависимости от источника используемой энергии можно разделить на три группы:
а) протекающие за счет энергии АТФ (это реакции карбоксилирования пропионил-КоА, пирувата, ацетил-КоА, р-метил-кро-тонил-КоА, геранил-КоА, мочевины и образования карбомилсрос-фата). Здесь фиксации углекислоты предшествует ее активация, которая осуществляется путем взаимодействия ее с АТФ. Важную роль в данном вопросе отводят биотину, который является простетической группой почти всех карбоксилаз, катализирующих АТФ-зависимые реакции карбоксилирования.
Процесс карбоксилирования с участием биотина можно представить схемой:
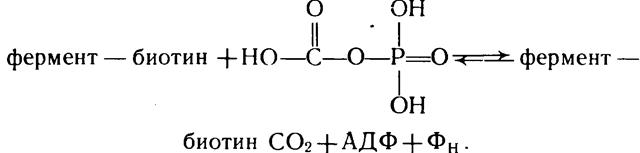
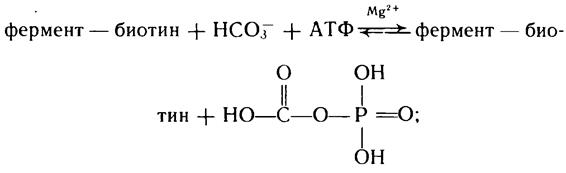
Образовавшийся комплекс фермент — биотин — СО2 способен передавать карбоксильную группу;
б) протекающие за счет энергии восстановленных форм пиридин нуклеотидов (это реакция карбоксилирования а-кетоглутарата, пирувата, рибулеза-5-фосфата, серина, глутамата, ацетата);
в) не требующие энергии извне (к этой группе реакций относятся: 5-аминоимидазолриботидкарбоксилазная, фосфоенолпируваткарбоксилазная, фосфоенилпируваткарбоксилазная и фосфоенолпируваткарбокситрансфосфорилазная реакции).
Анализируя данные по фиксации С02 во всех трех типах реакций, можно указать ряд факторов, влияющих на этот процесс. Главными из них являются следующие:
1) вариации концентраций и величин соотношения основных форм углекислоты в тканях (НСОз~ и рСОг);
2) изменение концентраций определенных ионов металлов, гвляющихся активаторами или ингибиторами карбоксилаз;
3) изменение концентраций субстрактов, подвергающихся карбоксилированию;
4) изменение степени обеспеченности клеток источниками энергии и рядом других метаболитов-аллостерических эффекторов карбоксилаз;
5) изменение в тканях концентраций коферментов карбоксилаз.
Наиболее универсальным из всех указанных путей воздействия на фиксацию углекислоты является 1-й, поскольку во всех реакциях карбоксилирования рСОг или НСОз~ служат субстратами. Концентрация же субстрата в ферментативных реакциях может играть определенную роль в интенсивности процесса его превращения. В ряде опытов было показано, что в условиях нормального рН инкубационной среды интенсивность фиксации углекислоты в тканях in vitro увеличивается по мере увеличения ее концентрации. Таким образом, можно заключить, что интенсивность реакций карбоксилирования во многом определяется концентрацией и величиной соотношения рССЬ и НСОГ в тканях. В условиях постоянного рН существует возможность значительно усиливать интенсивность процессов карбоксилирования за счет повышения до определенного уровня концентрации углекислоты в тканях.
Многочисленными исследованиями установлено, что мета-, болическое значение углекислоты не ограничивается отмеченным выше. Оказалось, что при изменении уровня углекислоты в тканях изменяется активность целого ряда ферментов гликолиза, трикарбонового цикла и азотистого обмена, увеличивается образование некоторых гормонов и др.
6. Роль углекислоты в регуляции реакций цикла
т р и к а р б о но в ы х кислот (ЦТК)
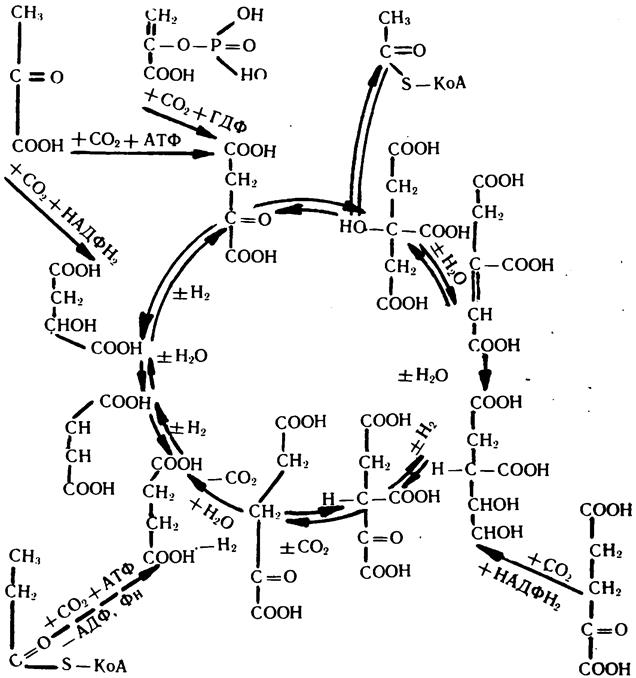
При рассмотрении роли углекислоты в регуляции реакций трикарбонового цикла следует отметить следующее. У многих биохимиков выработалось ошибочное представление о том, что цикл трикарбоновых кислот служит только механизмом окисления ацетил-КоА. Поэтому за ним закрепились такие названия, как «энергетическая мельница», «котел» и прочие, и почти во всех учебниках биохимии, а зачастую и в ряде монографий ЦТК представляют как последовательность реакций с указанием направленности процесса только по часовой стрелке. В связи с этим часто возникает вопрос: какова роль углекислоты в обеспечении нормального функционирования цикла? Ведь за один его оборот в реакциях декарбоксилирования образуется две молекулы СО2, а потребляется только одна СО2 при образовании оксалоацетата (карбоксилирование пирувата). Более того, так как в процессе оборота оксалоацетат снова регенерирует, то достаточно, чтобы раз образовалась молекула оксалоацетата, которая постоянно будет поддерживать цикл в функционирующем состоянии. Создается впечатление, что углекислота для функционирования цикла практически не нужна. Такие рассуждения в корне ошибочны. Ошибочность этих представлений заключается в следующем.
1. Почти все реакции ЦТК обратимы. В клетке они протекают в обоих направлениях. При обратном направлении (против часовой стрелки) углерод углекислоты будет не освобождаться, а использоваться в реакциях карбоксилирования для наращивания углеродного скелета ряда органических соединений и тогда углекислота должна поступать извне. В этом случае концентрация бикарбонатов и СО2 в клетке будет в значительной мере влиять на скорость этих реакций и соответственно на состояние и функцию ЦТК.
2. ЦТК принадлежит важная роль в обеспечении протекания не только окислительных, но и биосинтетических процессов, где он служит механизмом генерации многочисленных субстратов, идущих на синтез ряда аминокислот, азотистых оснований пури-новых и пиримидиновых нуклеотидов углеводов и липидов.
3. Значительное количество метаболитов ЦТК используется в процессах трансмембранного транспорта ряда анионов и пр. Существует мнение, что около 40% пула метаболитов ЦТК постоянно выключается из него и используется в различных биосинтетических процессах. Если это так, то нормальное функционирование ЦТК возможно только в тех случаях, когда не нарушены пути постоянной' образования его метаболитов. Так как 4 из 9 его метаболитов (ок-:алоацетат, малат, сукцинат, изоцитрат) образуются за счет карбоксилирования пирувата, фосфоенолпирувата, пропионил-КоА, ос-кетоглутарата, то функционирование цикла немыслимо без постоянного притока бикарбоната и углекислоты.
Подытоживая результаты различных опытов в этом направлении (Адлер и др., Остберг и др.), можно сказать, что активация ЦТК в значительной степени обусловливается как величиной рН биологических жидкостей, так и уровнем бикарбонатов и С02 в ней. Вместе с тем, в условиях постоянного рН биологических жидкостей активность ЦТК также увеличивается, но по мере увеличения в них концентрации углекислоты. Одним из подтверждений важности углекислоты и процессов ее фиксации в регуляции процессов окисления органических веществ в клетке являются результаты опытов Берри, Куна и Вернера, которые показали, что при инкубации в течение 40 мин изолированных «клеток печени крыс в среде, содержащей бикарбонат, концентрация малага повышалась примерно в 7 раз по сравнению с концентрацией в. среде без бикарбоната (фосфатный буфер), хотя рН сред были одинаковы.
Таким образом можно заключить, что углекислота, являясь одним из основных конечных продуктов обменных реакций ЦТК, одновременно выступает одним из мощных его регуляторов. Результаты иссследований последних лет дают основания полагать, что механизм регуляторного влияния углекислоты на функционирование ЦТК заключается не только в том, что она является одним из компонентов реакций карбоксилирования или декарбоксили-рования. Дело в том, что активность ряда ферментов, которые не имеют отношения к процессам карбоксилирования и декарбо-ксилирования, при постоянном рН среды могут существенно изменяться в зависимости от изменения в ней концентрации НСОз" и СО2 (даже в пределах физиологических колебаний).
Кроме того, полученные данные показывают, что уровень углекислоты определяет не только интенсивность функционирования указанных реакций, но влияет и на уровень биосинтетических процессов в организме, которые также находятся в неразрывной зависимости от функционирования трикарбонового цикла. К таким процессам относятся процессы обмена липидов, углеводов, белков, нуклеотидов и нуклеиновых кислот.
7. Роль углекислоты в процессах биосинтеза
Биосинтез аминокислот. Синтез таких аминокислот, как глицин, серии, цитрулин, глутаминовая и аспарагиновая кислоты и соответственно их амидов — глутамина и аспарагина фактически полностью зависит от интенсивности карбоксилирования соответствующих субстратов:
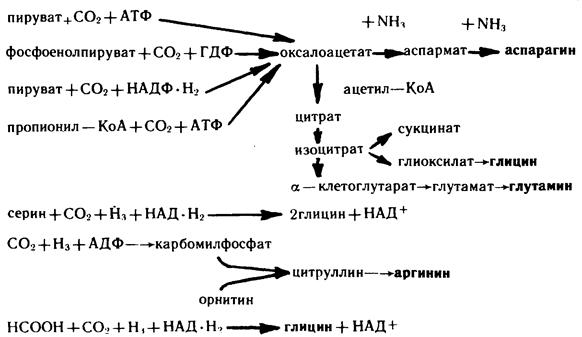
Биосинтез азотистых оснований нуклеотидов. Биосинтез пуриновых нуклеотидов. Первые важные данные о биосинтезе пуриновых оснований получены Бьюкененом, который скармливал животным различные меченые предшественники и определял места включения меченых атомов в пуриновое кольцо. Такие эксперименты были выполнены на птицах, у которых азот выделяется главным образом в виде мочевой кислоты, представляющей собой производное пурина. Оказалось, что два атома азота N (3, 9) пуринового кольца происходят из амидной группы глутамина, третий атом азота (1)—из аспарагиновой кислоты и, наконец, последний (7) — из глицина.
|
|
|
|
|
Дата добавления: 2017-01-14; Просмотров: 278; Нарушение авторских прав?; Мы поможем в написании вашей работы!