КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Напряжение, возникающее при суперспирализации ДНК, позволяет осуществлять воздействие на
|
|
|
|
расстоянии [32]
Неизвестно, как в хромосоме осуществляется контроль на большом расстоянии, В настоящее время описано несколько способов передачи
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
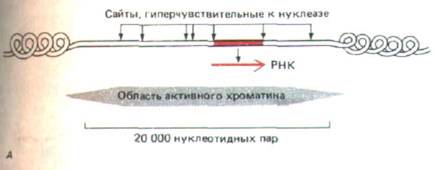
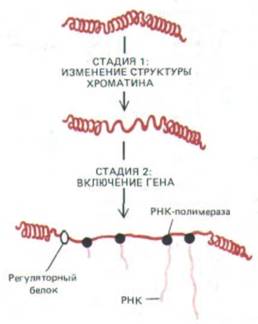
Рис. 10-40. Изменения хроматина в активных генах. А. Структура хроматина гена лизоцима в яйцеводе курицы, где этот ген активен. Как показано, область деконденсированного активного хроматина (определяемая по чувствительности к ДНКазе) содержит 24 000 нуклеотидных пар.
Она включает семь гиперчувствительных к нуклеазе сайтов и представляет собой специфическую последовательность ДНК, где, как полагают, нуклеосома заменена другими ДНК-связывающими белками. Б. Две стадии активации эукариотических генов. На стадии 1 структура небольшой области хроматина модифицирована таким образом, чтобы в ходе подготовки к транскрипции она могла деконденсироваться. На стадии 2 белки-
регуляторы связываются с определенными сайтами на изменившемся хроматине, чтобы индуцировать синтез РНК. Транскрипция у прокариот, по-
видимому, начинается со стадии 2 процесса контроля эукарнотических генов. (А А.Е. Sipple et al. In: Structure and Function of Eucaryotic Chromosomes [W. Hennig ed.]. Berlin: Springer Verlag, 1987.)
сигнала из одного участка молекулы ДНК в другой, удаленный от него на расстояние многих тысяч нуклеотидов. В одном случае в закрытой петле двойной спирали ДНК происходят топологические изменения, которые приводят к образованию суперскрученной ДНК. Суперспирализация ДНК лучше всего изучена на примере небольших кольцевых молекул, таких, как плазмиды и хромосомы некоторых вирусов (см. рис. 5-71). Однако такие же события могут произойти и с любым другим участком ДНК, ограниченным двумя концами, которые не могут свободно вращаться (например, с петлей хроматина, прочно связанной у основания).
|
|
|
Простой метод визуализации топологических напряжений, приводящих к суперспирализации ДНК, изображен на рис. 10-41. В двойной спирали ДНК с двумя фиксированными концами для компенсации каждых 10 открытых (не скрученных) нуклеотидных пар образуется один супервиток ДНК. Образование супервитка восстанавливает нормальный поворот спирали в остальных областях, где основания остаются спаренными; в ином случае (для схемы, приведенной на рис. 10-41, Б) потребовалось бы увеличить число пар, приходящихся на один виток, с 10 до 11. Спираль ДНК сопротивляется подобным деформациям, предпочитая ослаблять напряжение, изгибаясь в суперспирализованные петли.
У бактерий, таких, например, как E. соli, обнаружен особый фермент, называемый ДНК-гнразой, который, используя энергию гидролиза АТР, непрерывно образует в ДНК супервитки и тем самым поддерживает в петельных доменах постоянные механические напряжения. Это так называемые отрицательные супервитки: они закручены в противоположную сторону по сравнению с положительными супервитками, обра-
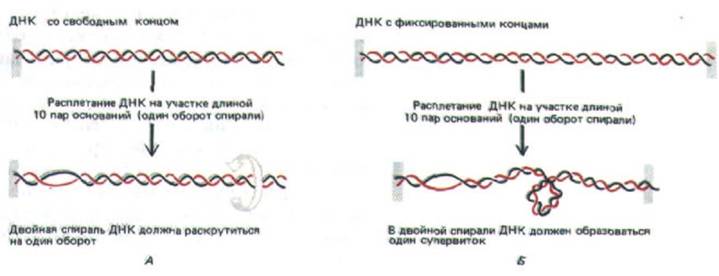
Рис. 10-41. Напряжение, возникающее при сверхспирализации ДНК, приводит к суперскручиванию молекулы. А. У молекулы ДНК с одним свободным концом (либо разрывом в одной из цепей, который служит шарниром) двойная цепь раскручивается на один виток на каждые 10
неспаренных нуклеотидных пар. Б. Если вращению что-либо препятствует, сверхвитки вводятся в ДНК путем раскручивания спирали в другом месте. В результате на каждые 10 неспаренных нуклеотидных пар возникает один сверхвиток. На этом рисунке изображена положительная сверхспираль (см. рис. 10-42).
|
|
|
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
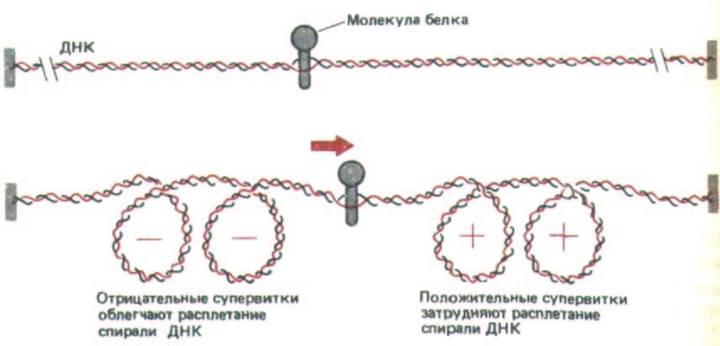
Рис. 10-42. Суперскручивание фрагмента ДНК с помощью белка, передвигающегося вдоль двойной спирали ДНК. Как и на рис. 10-41, Б, два конца ДНК зафиксированы, кроме того, принимается, что молекула белка закреплена относительно этих концов или же, что силы трения при движении затрудняют свободное вращение этого белка. Таким образом, при движении возникает избыток витков спирали, которые накапливаются в ДНК перед белком, а за ним образуется нехватка витков. Экспериментальные данные свидетельствуют о том, что продвижение молекул РНК-
полимеразы вдоль цепи ДНК сопровождается такими же сжатиями. Напряжение, возникающее в результате положительного суперскручивания впереди молекулы, приводит к тому, что этот участок ДНК труднее открыть, однако имеющееся напряжение должно облегчать сбрасывание ДНК с нуклеосом, что необходимо для работы полнмеразы при транскрипции хроматина (см. рис. 9-32).
зующимися при открытии фрагмента спирали. В связи с тем, что напряжение, вызываемое суперспирализацией, при этом снижается, расплетание двойной спирали ДНК у Е. coli энергетически более выгодно, чем расплетание ДНК, которая не подверглась суперспирализации.
В эукариотических клетках ДНК-гираза не обнаружена, а эукариотические ДНК топоизомеразы типов I и II снимают напряжение, вызванное суперспирализацией, а не усиливают его (см. разд. 5.3.10). Вот почему большая часть ДНК в эукариотических клетках не напряжена, Тем не менее при инициации транскрипции ДНК происходит раскручивание спирали (см. рис. 9-65). Более того, продвижение молекулы РНК-
полимеразы (а также других белков) вдоль ДНК приводит к появлению в ее молекуле положительного напряжения перед ферментом и отрицательного за ним (рис. 10-42). Вследствие подобного топологического изменения, событие, происшедшее в единственном сайте ДНК, может привести к возникновению сил, действующих по всей петле хроматина. Остается неясным, приводит ли подобное воздействие к запуску дальнейших событий, что было бы необходимо, если бы топологические изменения действительно имели отношение к контролю экспрессии у эукариотических генов.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2633; Нарушение авторских прав?; Мы поможем в написании вашей работы!