КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Метилирование ДНК у позвоночных способствует тому, что клетка придерживается выбранного пути
|
|
|
|
10-20
10-5
10-20
10.3.16. При делении клеток позвоночных тип метилирования ДНК передается по наследству [34]
Основания в молекуле ДНК в некоторых случаях подвергаются модификации. Например, мы уже говорили о том, что метилирование А в последовательности GATC приводит к ошибкам копирования в ходе репликации у бактерий (см. разд. 5.3.8.); напротив, метилирование А или С в определенном сайте защищает бактерию от воздействия ее собственных рестриктаз (см. разд. 4.6.2). В ДНК позвоночных содержится 5-
метнлцнтозин (5-метнл-С), который, по всей вероятности, не оказывает влияния на спаривание оснований (рис. 10-44,Л). Метилирование ограничено основанием С в последовательности CG. В связи с тем, что эта последовательность спарена точно с такой же последовательностью (но в обратной ориентации) на другой цепи ДНК, передача по наследству существующего типа метилирования ДНК обеспечивается простым механизмом копирования. Фермент, называемый поддерживающей метилазой, действует лишь на те последовательности CG, которые спарены с уже метилированными последовательностями CG. В результате существовавший ранее тип метилирования автоматически наследуется в ходе репликации ДНК (рис. 10-45).
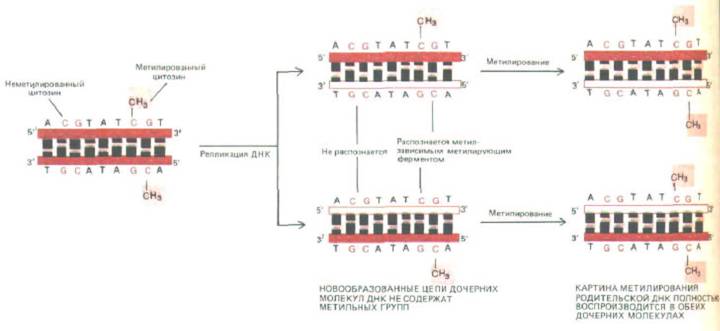
Рис. 10-45. Точное наследование порядка метилирования ДНК. В ДНК позвоночных метилирована большая часть остатков цитозина, входящих в состав последовательности CG (см. рис. 10-44). После того как разметка ДНК метильными группами проведена, каждый сайт метилирования наследуется дочерними молекулами ДНК. Это означает, что изменение в метилированой разметке ДНК сохраняется и наследуется в клетку.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
Некоторые рестриктазы разрезают ДНК в области, содержащей динуклеотид CG, и для их действия необходимо, чтобы эта последовательность не была метилирована. Например, Hpall разрезает последовательность CCGG, но не способна это сделать, если средний С метилирован. Таким образом, чувствительность ДНК к ресгриктязс Hpall может служить тестом на метилирование определенных сайтов. Фермент Нра..-метилаза в обычных условиях защищает любую бактерию от своей собственной рестрикгазы Hpall. Следовательно, эту метилазу можно использовать для введения оснований 5-метил-С в определенные последовательности CG (а именно в CCGG) в составе клонированной молекулы ДНК (при клонировании ДНК в E.coli метилирование по CG-сайтам утрачивается). С помощью этой методики удалось показать, что каждая отдельная метилированная последовательность CG обычно сохраняется в ходе многих клеточных делений в культуре клеток позвоночных, а неметилированные последовательности CG так и остаются неметилированными.
Автоматическое наследование 5-метил-С ставит проблему «курицы и яйца»: на какой стадии происходит первое метилирование у позвоночных? Установлено, что при введении неметилированной ДНК в оплодотворенную яйцеклетку мыши почти все CG-сайты этой ДНК метилируются (хотя существует одно важное исключение, описанное ниже). Таким образом, большая часть генома млекопитающих может оказаться сильно метилированной. Поскольку известно, что в норме поддерживающая метилаза не способна метилировать ДНК, лишенную метальных групп, остается предположить, что в яйцеклетке должен присутствовать другой фермент: учреждающая метилаза. Вероятно, выполнив свою функцию, она исчезает, а за сохранение метилированных нуклеотидов в ДНК клеток развивающихся тканей отвечает уже другая, поддерживающая метилаза.
|
|
|
развития [35]
Какова роль метилирования CG? Эксперименты, проведенные с использованием рестриктазы Hpall, свидетельствуют о том, что ДНК неактивных генов метилирована сильнее, нежели ДНК активных генов. Более того, оказалось, что неактивный ген, ДНК которого метилирована, после активации теряет большую часть своих метальных групп. Убедительное доказательство того, что именно метилирование влияет на экспрессию генов, получено в опытах с 5-азацитозином (5-aza-C, см. рис. 10-44,6), который на короткий период добавляли к культивируемым клеткам. 5-aza-C, аналог основания, не способный метилироваться, включается в состав ДНК и действует в качестве ингибитора поддерживающей метилазы, снижая таким образом общий уровень метилирования ДНК. В клетках, обработанных 5-aza-C, определенные ранее неактивные гены активировались и одновременно получали неметилированные остатки С. После разовой активации активное состояние этих генов поддерживалось и в отсутствие 5-aza-C во многих последующих поколениях клеток. Этот факт указывает на то, что изначальное метилирование генов способствовало их пребыванию в неактивном состоянии.
На основании результатов, полученных в опытах с 5-азацитозином, было выдвинуто предположение, что метилирование ДНК может играть главную роль в возникновении различных типов клеток. Это означало бы, что специализация клеток у позвоночных и беспозвоночных осу-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
ществляется разными путями. Последующее изучение показало, однако, что роль метилирования ДНК в дифференцировке клеток, по-видимому, не столь существенна и носит скорее вспомогательный характер. Важные переключения в ходе развития осуществляются регуляторными белками, способными влиять на активность генов вне зависимости от их метилирования. Например, Х-хромосома самок сперва конденсируется и инактивируется, и лишь затем некоторые из ее генов метилируются сильнее. И наоборот, определенные гены, активные в печени, в ходе развития включаются тогда, когда они еще полностью метилированы, и лишь позднее уровень их метилирования начинает снижаться.
|
|
|
Значение метилирования ДНК для экспрессии генов в значительной степени прояснилось в результате опытов по трансфекции.
Например, тканеспецифичный ген, кодирующий актин мышц, выделяли как в полностью метилированной, так и в полностью неметилированной формах. При введении двух модификаций этого гена в культуру мышечных клеток оба варианта транскрибировались одинаково эффективно. Если же этот ген вводили в фибробласты, где он в норме не экспрессируется, неметилированный вариант транскрибировался на низком уровне, который тем не менее был выше, чем у введенного метилированного гена или у эндогенного гена, присутствующего в фибробласте и также метилированного. На основании этих опытов можно сделать вывод, что у позвоночных метилирование ДНК используется для закрепления пути развития, выбранного иными способами.
В отдельных случаях можно с достаточной точностью определить, могут ли последовательности ДНК, транскрибируемые с высокой эффективностью в клетках одного типа, транскрибироваться в клетках позвоночных другого типа. Подобные эксперименты показали, что уровни транскрипции в двух типах клеток позвоночных иногда различаются более чем в 106 раз. Неэкспрессируемые гены позвоночных гораздо менее «расплывчаты» в отношении транскрипции, чем неэкс-прессируемые гены бактерий, у которых наибольшие известные различия в скорости транскрипции экспрессируемых и неэкспрессируемых генов не превышают 1000-кратный уровень. Способствуя дальнейшему снижению транскрипции генов, включенных иным способом, метилирование ДНК, по-видимому, вносит свой вклад в это различие.
10.3.18. CG-богатые островки позволяют выявить у млекопитающих около 30000 генов «домашнего хозяйства» [36]
В ходе эволюции метилированные остатки в геноме имеют тенденцию к исчезновению, что объясняется специфическими особенностями механизма репарации. Дело в том, что случайное дезаминирование неметилированного цитозина приводит к возникновению урацила, который в норме не входит в состав ДНК и потому легко распознается ферментом репарации, вырезается и затем заменяется на цитозин. Последстви.
|
|
|
случайного дезаминирования остатка 5-метилцитозина не могут быть устранены подобным образом, так как при дезаминировании 5-метил-С образуется тимин, который неотличим от других, немутантных остатков тимина, входящих в состав ДНК. Следовательно, в ходе эволюцш метилированные остатки цитозина в геноме имеют тенденцию превращаться в тимин.
После того, как около 400 млн. лет назад произошла дивергенци. позвоночных и беспозвоночных, подобным образом утерялось более трех из каждых четырех CG, что привело к значительному уменьшению содержания этого динуклеотида в геноме позвоночных. Оставшиеся
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
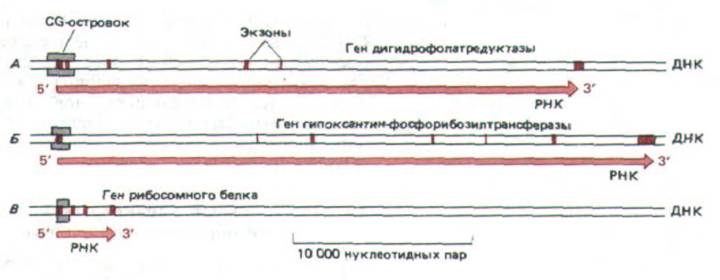
Рис. 10-46. CG-островки в трех генах «домашнего хозяйства» млекопитающих. Прямоугольниками отмечен размер каждого островка, которые, как показано, окружают промоторы каждого гена. Следует отметить, что у большинства генов млекопитающих экзоны (выделены цветом)
короче, нежели интроны. (По А. Р. Bird, Trends Genet. 3: 342-347, 1987.)
последовательности CG очень неравномерно распределены по геному: существуют отдельные области длиной 1000-2000 нуклеотидов (СG-
островки), в которых содержание CG в 10-20 раз выше, чем в среднем по геному. Подобные островки окружают промоторы так называемых генов «домашнего хозяйства», т.е. тех генов, которые кодируют белки, необходимые для жизнедеятельности клетки и, следовательно, экспрессируемые в клетках любого типа (рис. 10-46). Эти гены можно противопоставить тканеспецифичным генам, кодирующим белки, присущие лишь строго определенным типам клеток.
Распределение CG-островков можно легко объяснить как побочный эффект развития системы метилирования CG, предназначенный для снижения экспрессии неактивных генов у позвоночных (рис. 10-47). В половых клетках все тканеспецифичные гены (за исключением тех, которые присущи яйцеклетке и спермию) неактивны и метилированы. За долгую эволюцию их метилированные CG были утеряны при случайном дезаминировании. Однако CG-последовательности в промоторных областях генов, активных в половых клетках (включая все гены «домашнего хозяйства»), оставались деметилированными и при спонтанном дезаминировании всегда репарировались. Полагают, что эти гены узнаются сайт-
специфическими ДНК-связывающими белками, которые присутствуют в половых клетках и удаляют любые метильные группы в области промоторов генов «домашнего хозяйства». Эксперименты с клонированными генами показывают, что лишь CG, входящие в состав CG-островков, остаются неметилированными, если в яйцеклетку мыши инъецировать совершенно неметилированную ДНК.
Известно, что геном млекопитающих (размером около 3 х 109 нуклеотидных пар) содержит около 30 000 CG-островков, каждый из которых имеет в длину около 1000 нуклеотидных пар. Большинство островков совпадает с 5'-концом транскрипционной единицы и таким образом,
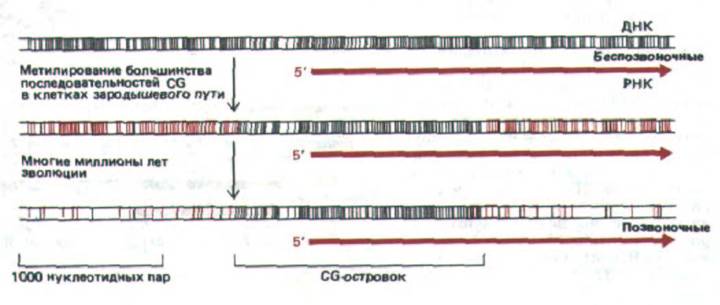
Рис. 10-47. Схема, иллюстрирующая заметную нехватку последовательностей CG и наличие CG-островков. Черными линиями отмечено расположение неметилированных динуклеотидов CG в последовательности ДНК, а красными линиями обозначены метилированные CG-
динуклеотиды. Метилированные гены беспозвоночных неизвестны.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
по-видимому, с 5'-концом гена. В связи с тем, что ДНК, окружающую CG-островки, можно выделить и клонировать отдельно, относительно просто решается задача идентификации и характеристики жизненно важных генов (генов «домашнего хозяйства»). Вероятно, несколько десятков тысяч остальных генов являются тканеспецифичными и не экспрессируются в половых клетках. Так как большая часть их последовательностей CG утеряна, эти тканеспецифичные гены гораздо труднее выявить в геноме.
10.3.19. Сложный характер регуляции генов необходим для образования многоклеточного организма [37]
Клетки эмбриона имеют много общего с компьютером, поскольку постоянно получают информацию о своем расположении в данный момент и объединяют эту информацию с поступившей ранее для того, чтобы на каждой стадии развития действовать соответствующим образом.
Изучение дрозофилы генетическими методами показало, что в образовании и поддержании основного плана строения тела участвует относительно небольшое число (порядка 100) генов, кодирующих главные регуляторные белки, взаимодействующие между собой. В любом многоклеточном организме подавляющее большинство генов (и жизненно важных, и тканеспецифичных), вероятно, регулируются посредством сложных контролирующих цепочек, исходящих от генов главных регуляторных белков. Если в регуляции генов эукариот центральную роль играют механизмы, сильно отличающиеся от бактериальных (например, механизмы, зависящие от прямого наследования структуры хроматина), можно ожидать, что именно эти механизмы контролируют некоторые гены главных белков-регуляторов.
В настоящее время ничего неизвестно о том, как осуществляется контроль за действием генов главных белков-регуляторов у позвоночных, однако начали появляться первые данные о том, как эти гены контролируются у дрозофилы. Например, гомеотические гены, определяющие разное развитие отдельных сегментов тела мухи, находятся в двух сложных локусах Antennapedia и Bithorax. Комплекс биторакса, ответственный за дифференцировку двух грудных сегментов и восьми сегментов брюшка, содержит три транскрипционные единицы, называемые Ubx, abd-A и abd-B. Каждая транскрипционная единица, по-видимому, кодирует семейство белков-регуляторов, образующихся в результате альтернативного сплайсинга РНК. Полагают, что кодирующие
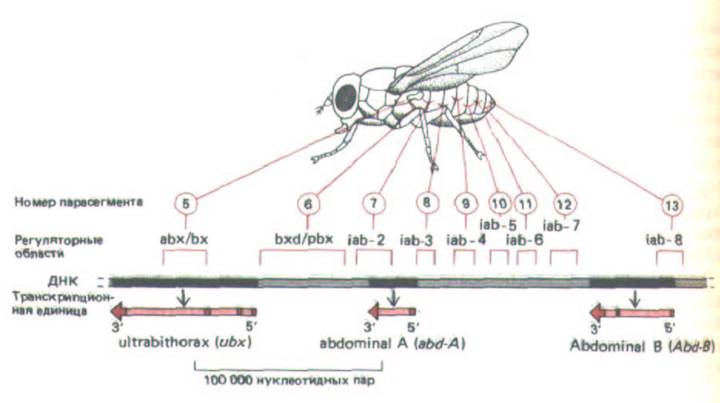
Рис. 10-48. Организация биторального комплекса у дрозофилы. В этой важной области хромосомы размером 300 000 нуклеотидных пар содержатся три гена - Ubx, abd-A и Abd-B. которые кодируют главные регуляторные белки, контролирующие развитие груди и брюшка.
Гомеотические мутации помогли выявить еще девять групп регуляторных последовательностей ДНК. Каждая из этих групп необходима для развития указанных парасегментов, а также других парасегментов, расположенных ближе к концу тела. Считается, что эти регуляторные последовательности ДНК действуют как энхансеры, контролируя экспрессию одного из близлежащих генов, а порядок их расположения на молекуле ДНК соответствует расположению тех сегментов тела, на которые они воздействуют (см. рис. 10-49). (По М. Peifer, F. Karch and W.
Bender, Genes Devel. 1: 891-898, 1987.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
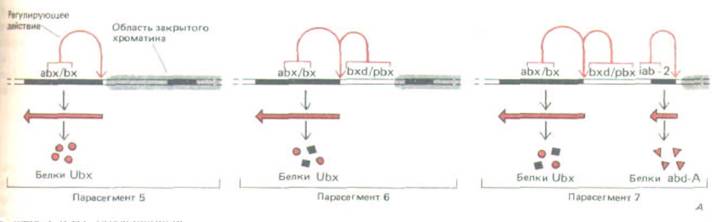
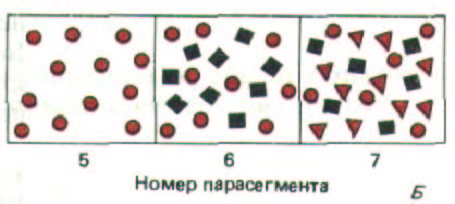
Рис. 10-49. Схема, объясняющая тачное соответствие между положением в хромосоме каждого регулирующего участка биторального гомплекса и расположением на теле мухи тех парасегментов, на которые оказывают влияние мутации в этой области. А. Контроль на уровне изменения структуры хроматина. Предполагается, что хроматин деконденсируется или же постепенно активируется во все более отдаленных парасегментах, таким образом, в парасегменте 5 открыта только регуляторная область вbх/bх, тогда как все остальные регуляторные участки экспонированы в самых удаленных парасегментах. находящихся под контролем того комплекса (парасегмент 13, см. рис. 10-48). На данном рисунке изображены лишь три парасегмента. Ген Ubx может образовывать несколько различных транскриптов (обозначенных здесь квадратиками и кружочками), отбор которых контролируется регуляторными областями. Таким образом, биторальный комплекс образует в каждом из парасегментов свою смесь белков-регуляторов (Б).
области этого комплекса содержат менее 20 000 нуклеотидных пар, тогда как регуляторные последовательности насчитывают приблизительно 300000 нуклеотидных пар. По-видимому, регуляторные области состоят из набора энхансеров, расположение которых по длине хромосомы соответствует расположению сегментов тела, подвергающихся действию этих энхансеров (рис. 10-48). Эти факты в совокупности с данными о строении тела мутантных мух, у которых один из энхансеров транслоцирован из одной части комплекса биторакса в другую, позволяют выдвинуть предположение о регуляции генов внутри комплекса биторакса на основе изменений в структуре хроматина. Согласно этому предположению, в клетках, образующих комплекс биторакса и расположенных все ближе к концу тела, последовательно открываются лежащие один за другим домены хроматина. Это дает возможность энхансерам, локализованным в данных доменах, активироваться в строгой очередности (рис. 10-49).
Весьма вероятно, что механизмы, контролирующие гены белков-регуляторов, не менее сложны.
Заключение
В организме животных и растений функционируют механизмы, ответственные за то, что в разных клетках транскрибируются
разные гены. Так как многие специализированные клетки обладают способностью поддерживать свои уникальные свойства при выращивании их в
культуре, механизмы регуляции генов должны быть стабильными и наследуемыми. Прокариоты и дрожжи представляют собой весьма удобную
модельную систему, с помощью которой можно изучать механизмы регуляции генов. Некоторые из этих механизмов могут также принимать
участие в возникновении специализированных типов клеток у высших эукариот. Один из них - конкурентное взаимодействие между двумя или
более главными белками-регуляторами, каждый из которых, подавляя образование другого, стимулирует свой собственный синтез.
Изучение экспрессии генов, сконструированных методами генной инженерии, а также фрагментов, встроившихся в случайные сайты
генома трансгенных животных,, показало, что большая часть генов высших эукариот контролируется смесью диффундирующих белков-
регуляторов, уникальных для каждого типа клеток. Кроме того, на экспрессию генов у высших эукариот может влиять переход хроматина
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
в более или менее конденсированное состояние; у позвоночных транскрипцию неактивных генов подавляет метилирование. Однако для
большинства генов эти дополнительные уровни контроля могут либо определяться, либо преодолеваться диффундирующими белками-
регуляторами. До сих пор неизвестно, каким образом контролируются гены, кодирующие главные белки-регуляторы, которые, собственно, и
детерминируют у позвоночных тип клетки.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 811; Нарушение авторских прав?; Мы поможем в написании вашей работы!