КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Микротрубочки определяют местоположение эндоплазматического ретнкулума и аппарата Гольджи
внутри клетки [50]
Двигательные белки, сходные с кинезином и динеином, существуют не только в нейронах. По-видимому, они имеются во всех клетках, где есть микротрубочки, и ряд важных организующих функций микротрубочек осуществляется, судя по всему, именно благодаря этим белкам. Как показали недавние исследования in vitro, кинезин прикрепляется к мембране эндоплазматического ретикулума и может растягивать ее вдоль ориентированных микротрубочек, превращая эту органеллу в характерную сложную сеть. Работы, проведенные на интактных клетках, тоже указывают на то, что ретикулум растягивается по ходу микротрубочек в направлении от центросомы, как и следует ожидать, если это происходит при участии кинезина. Наконец, иммунофлуоресцентными методами показано, что тонкие края цистерн ретикулума в периферических участках клеток часто располагаются вдоль микротрубочек (рис. 11-72, А и Б).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
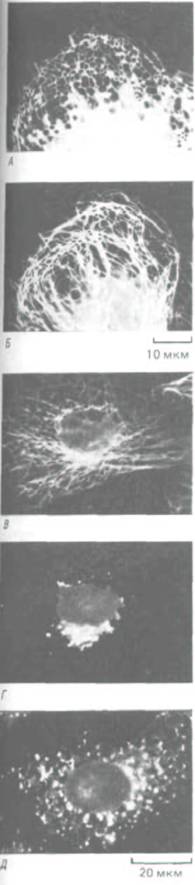
Рис. 11-72. Расположение мембран эндоплазматического ретикулума (ЭР) и микротрубочек в культивируемых клетках. А.
Иммунофлуоресцентное окрашивание белков ЭР выявляет цистерны ЭР в виде кружевной сети на периферии клеток. Б. Расположение микротрубочек в той же клетке. В, Г и Д. Влияние микротрубочек на аппарат Гольджи. Если в культивируемой клетке одновременно окрасить флуоресцентными антителами и микротрубочки (В), и мембраны Гольджи (Г), последние обнаружатся в виде скопления вокруг центросомы. Но если клетку обработать нокодазолом, вызывающим деполимеризацию микротрубочек, то мембраны Гольджи будут распределены по всей цитоплазме (Д), как это бывает во время митоза. [С любезного разрешения Mark Terasaki и Lan Во Chen (A, Б); Viki Allan и Thomas Kreis (В, Г. Д).]
В то время как мембрана эндоплазматического ретикулума активно перемещается в сторону от центросоми по «дорожкам» из микротрубочек (оставаясь соединенной на другом конце с ядерной оболочкой), мембраны цистерн Гольджи, по-видимому, транспортируются в обратном направлении, как если бы они были связаны с динеиноподобными белками; в результате аппарат Гольджи оказывается вблизи центросомы (рис. 11-72,5 и Г). И аппарат Гольджи, и эндоплазматический ретикулум во время митоза подвергаются сильной фрагментации (разд.
13.5.16), и когда потом в цитоплазме вновь образуются микротрубочки, именно их ориентация, вероятно, направляет восстановление этих органелл из мелких пузырьков и фрагментов мембран (рис. 11-72, Д).
Заключение
Микротрубочки образуются путем полимеризации молекул тубулина, после чего эти молекулы гидролизуют прочно связанный с ними
GTP (этот процесс несколько отстает от полимеризации). Микротрубочки растут медленно, нестабильны и склонны к взрывообразной, «катастрофической» деполимеризации: однако они могут стабилизироваться при ассоциации с другими структурами, которые прикрывают
(«кэпируют») их концы. Центры организации микротрубочек, такие как центросомы, все время инициируют образование новых микротрубочек, которые растут в случайных направлениях. Любая микротрубочка, которая натолкнется на какую-либо структуру, способную кэпировать
свободный плюс-конец этой микротрубочки, будет избирательно стабилизирована, тогда как другие со временем деполимеризуются. Полагают, что именно этот процесс в основном определяет полярность и расположение систем микротрубочек в клетке.
В тех микротрубочках, которые образовались в нужных местах, субъединицы тубулина подвергаются модификации - ацетилированию
и детирозилированию. Эти модификации играют роль «маркеров» зрелых микротрубочек и создают участки для связывания специальных белков, ассоциированных с микротрубочками (БАМ), которые еще больше повышают устойчивость микротрубочек к деполимеризации и адаптируют их
для выполнения специфических функций в клетке. Особая группа таких белков использует энергию гидролиза АТР для однонаправленного
перемещения вдоль по микротрубочке, обеспечивая этим направленное движение в цитоплазме клеточных органелл и их правильную
пространственную организацию.
11.5. Промежуточные филаменты [51]
Промежуточные филаменты (ПФ)-это жесткие и прочные белковые волокна в цитоплазме большинства клеток высших эукариот.
Структура их напоминает переплетенные канаты, а толщина составляет 8-10 нм,
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
т. е. промежуточная между толщиной толстых и тонких филаментов в мышцах, где ПФ были впервые описаны; по толщине они занимают также промежуточное место между актиновыми филаментами и микротрубочками. В большинстве животных клеток они формируют характерную «корзинку» вокруг ядра, откуда по слегка искривленным путям тянутся к периферии клетки. Особенно много ПФ там, где клети подвергаются механическим нагрузкам, например в эпителиях, где это нити участвуют в соединении клеток друг с другом (при помощн десмосом, см. разд.
14.1.4), в нервных волокнах и во всей цитоплазме гладкомышечных клеток. При экстракции клеток растворами с высокой и низкой ионной силой и солюбилизации неионными детергентами ПФ остаются целыми, тогда как большая часть остальных цитоскелетных структур разрушается.
Фактически термин «цитоскелет» был первоначально введен именно для обозначения этих чрезвычайно стойких и нерастворимых волокон.
11.5.1. Промежуточные филаменты образуются из фибриллярных полипептидов четырех типов [51, 52]
В отличие от мономеров актина и тубулина, которые представляют собой глобулярные белки, субъединицы ПФ имеют вытянутую, фибриллярную форму. Они объединяются в продольные пучки, где перекрываются по длине, так что образуют длинные нити с высокой механической прочностью. В латеральных взаимодействиях, за счет которых строятся ПФ, нередко участвует лишь часть молекулы белковой субъединицы ПФ, поэтому структура остальной ее части может значительно варьировать, не изменяя общего строения нити. В связи с этим ПФ в отличие от актиновых филаментов и микротрубочек построены из полипептидов с весьма различной молекулярной массой - от 40 до 130 тыс. в зависимости от типа клеток.
Промежуточные филаменты по их первичной структуре делят на четыре большие группы (табл. 11-5). Белки ПФ типа I наиболее характерны для эпителиальных клеток и включают два подсемейства керати-
| Таблица 11-5. Главные типы белков промежуточных филаментов | ||
| Образующий полипептид (мол. масса) | Локализация | |
| Тип I | Кислые кератины (40000-70000) Нейтральные и основные кератины (40 000-70 000) | Эпителиальные клетки и производные эпидермиса (волосы, ногти и т.п.) |
| Тип II | Виментин (53000) | Многие клетки мезенхимного происхождения; часто экспрес-сируется клетками в культуре |
| Мышечные клетки | ||
| Десмин (52000) Глиальный фибриллярный кислый белок (45000) | Глиальные клетки (астроциты и некоторые шванновские клетки) | |
| Тип III | Белки нейрофиламентов (около 1300001}, 1000001} и 60000) | Нейроны |
| Типы IV | Ядерные ламины А, В и С (65000 -75000) | Ядерная ламина во всех клетках |
| 1) Из-за того, что эти белки при электрофорезе в гелях с додецилсульфатом натрия мигрируют аномально медленно, раньше их молекулярную массу считали большей. |
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
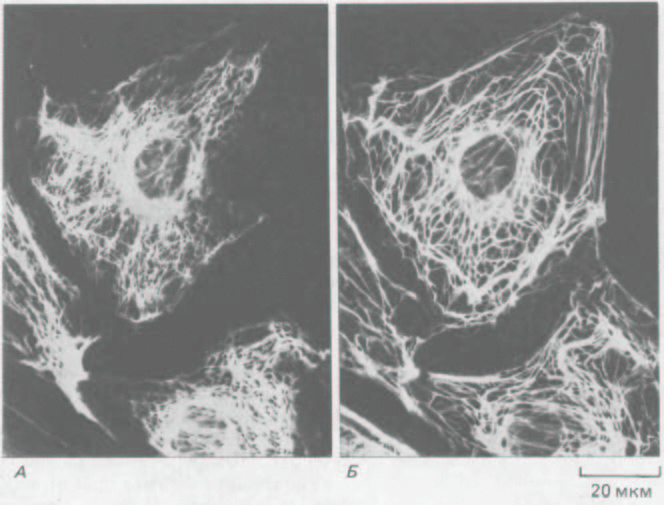
Рис. 11-73. Иммунофлуоресцентная микрофотография эпителиальных клеток кенгуровой крысы (РtК2) в интерфазе. Клетки окрашены одновременно антителами к виментину (А) и к кератину (Б). Обратите внимание, что содержащиеся в клетке системы виментиновых и кератиновых филаментов существуют раздельно, хотя и имеют сходное распределение. (С любезного разрешения Магу Osborn.)
нов: кислые кератины и нейтральные или основные кератины. Кератиновые филаменты - всегда гетерополимеры, образованные поровну субъединицами каждого из этих двух подсемейств. Вообще кератины - самая обширная группа белков ПФ; известно уже не менее 19 различных форм в составе эпителиев человека и еще 8 в волосах и ногтях. Многие типы эпителиев, различающихся морфологически и функционально, синтезируют также разные формы кератинов.
К белкам ПФ типа II относятся 1) виментин, 2) десмин и 3) глиальный фибриллярный кислый белок. Виментин широко распространен в клетках мезенхимного происхождения, включая фибробласты, клетки эндотелия кровеносных сосудов и лейкоциты; он часто образуется в культивируемых клетках и временно появляется в различных клетках в ходе онтогенеза. Десмин содержится в клетках мышц, как гладких, так и поперечнополосатых, а глиальный фибриллярный кислый белок образует глиальные филаменты в определенного рода клетках глии (астроцитах и некоторых шванновских клетках) в нервной системе. Все эти белки способны in vitro к самосборке с образованием гомополимеров, а также к образованию гетерополимеров с другими белками типа II. Последняя способность проявляется и in vivo: в клетках некоторых типов были обнаружены сополимеры виментина с десмином и виментина с глиальным фибриллярным кислым белком.
Из белков ПФ типа III построены нейрофиламенты - важный компонент цитоскелета в аксонах и дендритах нервных клеток. У позвоночных три таких белка, их называют «нейрофиламентным триплетом». И наконец, белки ПФ типа lV -это ядерные ламины (разд. 11.5.5);
они сходны с другими белками ПФ по аминокислотной последовательности, но имеют несколько характерных отличий. Наиболее примечательно то, что они образуют высокоупорядоченные двумерные сети из филаментов, подвергающиеся быстрой разборке и сборке на определенных стадиях митоза.
Все клетки эукариот синтезируют ядерные ламины и по крайней мере один тип цитоплазматических белков ПФ. В некоторых клетках есть цитоплазматические ПФ двух типов, образующие раздельные структу-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
ры. Таковы, например, некоторые эпителиальные клетки, содержащие отдельные системы кератиновых и виментиновых филаментов (рис, 11-73).
|
|
Дата добавления: 2015-08-31; Просмотров: 513; Нарушение авторских прав?; Мы поможем в написании вашей работы!