КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Разные клетки по-разному реагируют на один и тот же сигнал
|
|
|
|
12-4
12-3
Медиаторов и нейромедиаторов
Химические сигнальные механизмы различаются по расстояниям, на которых они действуют: 1) в случае эндокринной сигнализации
специализированные эндокринные клетки выделяют гормоны, которые разносятся кровью и воздействуют на клетки-мишени, находящиеся иногда в самых разных частях организма; 2) в случае паракринной сигнализации клетки выделяют локальные химические медиаторы, которые поглощаются, разрушаются или иммобилизуются так быстро, что успевают подействовать только на клетки ближайшего окружения, быть может, в радиусе около миллиметра; 3) при синаптической передаче, используемой только в нервной системе, клетки секретируют нейромедиаторы в специализированных межклеточных контактах, называемых химическими синапсами, Нейромедиаторы диффундируют через синаптическую щель, обычно на расстояние около 50 нм, и воздействуют только на одну постсинаптическую клетку-мишень (рис. 12-2). В каждом случае мишень реагирует на определенный внеклеточный сигнал с помощью специальных белков, называемых рецепторами, которые связывают сигнальную молекулу и инициируют ответ. Многие сигнальные молекулы и рецепторы используются в передаче сигнала и по эндокринному, и по паракринному, и по синаптическому типу. Главные различия касаются быстроты и избирательности воздействия сигнала на определенные мишени.
12.1.1. Эндокринные клетки и нервные клетки специализированы для разных типов химической сигнализации [1]
Эндокринные и нервные клетки совместно координируют разнообразные функции миллиардов клеток, из которых состоит тело у высших животных. Эндокринные клетки обычно собраны в специальные железы
|
|
|
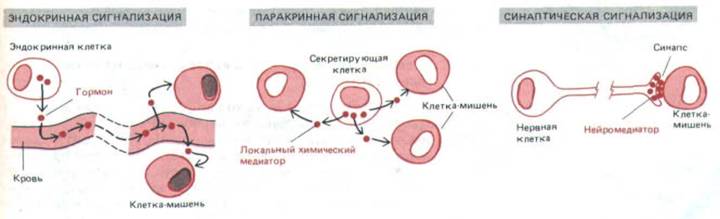
Рис. 12-2. Три формы сигнализации с помощью секретируемых молекул. Не все нейромедиаторы действуют в синапсах, как показано на рисунке; некоторые из них работают как локальные химические медиаторы (по паракринному типу), влияя сразу на целую группу соседних клеток-
мишеней.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
и выделяют свои гормоны во внеклеточную (интерстициальную) жидкость, окружающую все клетки в тканях. Отсюда молекулы диффундируют в капилляры и разносятся с кровью по всему телу. В каждой ткани они проникают из капилляров в интерстициальную жидкость и могут связываться клетками-мишенями. Поскольку распространение эндокринного сигнала определяется диффузией и кровотоком, оно происходит сравнительно медленно: обычно требуются минуты, чтобы гормон достиг своей мишени. Кроме того, специфичность сигналов в эндокринной системе всецело зависит от химической природы сигнального вещества и рецепторов на поверхности клетки-мишени: каждый тип эндокринной клетки секретирует в кровь свой гормон, и любая клетка, имеющая комплементарный рецептор для этого гормона, ответит реакцией, характерной для данной клетки (рис. 12-3, А).
Работа нервных клеток отличается гораздо большей быстротой и точностью. Они могут передавать информацию на большие расстояния по нервному волокну с помощью электрических импульсов со скоростью более 100 м/с. Только в нервных окончаниях, где высвобождается нейромедиатор, эти импульсы преобразуются в химические сигналы. Химический сигнал нервной клетки может действовать как паракринный или как синаптический. В первом случае нейромедиатор, подобно локальному химическому медиатору, диффундирует наружу и влияет на все соседние клетки-мишени, у которых имеется надлежащий рецептор. При синаптической передаче сигнал гораздо более точен и действие нейромедиатора ограничено единственной клеткой-мишенью, даже если соседние клетки имеют рецепторы для того же нейромедиатора (рис. 12-3, Б). Поскольку расстояние, на которое нейромедиатор должен в таких случаях диффундировать, меньше 100 нм, процесс длится менее миллисекунды (рис. 12-2).
|
|
|
Гормоны в крови и интерстициальной жидкости очень сильно разбавляются, и поэтому они должны быть способны действовать в чрезвычайно низких концентрациях (обычно менее 10-8 М); разбавление же нейромедиаторов на их коротком пути незначительно, и их концентрация около мембраны постсинаптической клетки может быть сравнительно высокой. Например, концентрация ацетилхолина в синаптической щели нервно-мышечного соединения составляет около 5-10-4 М. В соответствии с этим рецепторы нейромедиатора в синапсе обладают относительно низким сродством к своему лиганду и не могут заметным образом реагировать на низкие концентрации нейромедиатора, приходящего путем диффузии от соседних синапсов. Нейромедиатор быстро удаляется из синаптической щели специальными гидролитическими ферментами или мембранными транспортными белками, которые перекачивают его обратно в нервное окончание. Этим достигается точность воздействия сигнала не только в пространстве, но и во времени: короткое, «импульсное» освобождение нейромедиатора вызывает быстрый и краткий ответ, что позволяет сохранять временные характеристики сигнала при передаче его от клетки к клетке.
12.1.2. Главным регулятором эндокринной системы служит гипоталамус [1, 2]
В определенном участке мозга- гипоталамусе -эндокринная и нервная системы физически и функционально связаны друг с другом.
Гипоталамус расположен непосредственно над гипофизом, с которым соединен ножкой гипофиза. Гипоталамус выполняет свою связующую роль с помощью клеток, сочетающих особенности нейрона и эндокринной клетки; соответственно их называют нейроэндокринными (нейросекреторными)
клетками. Большая часть таких клеток гипоталамуса отвечает на
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
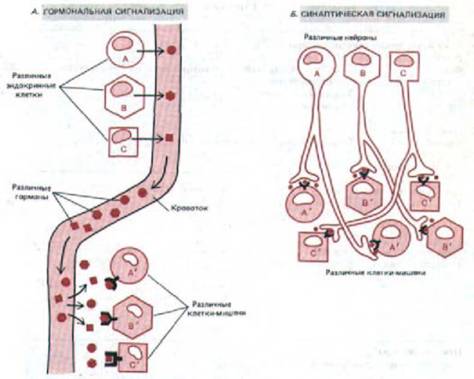
Рис. 12-3. Различие между гормональной (А) и синаптической (Б} передачей сигналов. Эндокринные клетки выделяют в кровь множество гормонов, и клетки-мишени, чувствительные к данному гормону, т.е. имеющие рецепторы для его связывания, «вылавливают» соответствующий гормон из внеклеточной жидкости. При синаптической же передаче специфичность определяется тесным контактом между окончанием нервного волокна и той клеткой-мишенью, которой это волокно передает сигнал: медиатор, выделяемый нервным окончанием, достигает только этой клетки.
Для специфической коммуникации с различными клетками-мишенями разные эндокринные клетки должны использовать разные гормоны, в то время как многие нервные клетки могут использовать один и тот же нейромедиатор, сохраняя при этом специфичность воздействия.
стимуляцию их нейронами других отделов мозга, выделяя в кровеносные сосуды гипофизарной ножки определенный пептидный гормон, который затем специфически стимулирует или подавляет секрецию гипофизом другого гормона. (Другие нейроэндокринные клетки гипоталамуса посылают аксоны в гипофиз и через них выделяют свои секреты
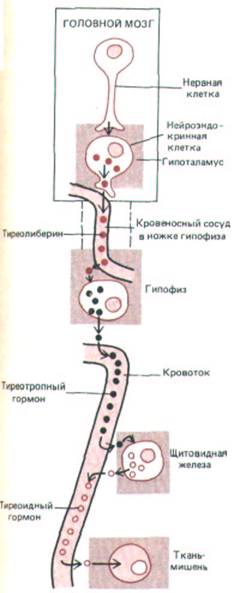
Рис. 12-4. Секреция тиреоидного гормона опосредованно регулируется нервной системой. Определенные нейросекреторные клетки гипоталамуса при стимуляции их нейронами высших отделов мозга выделяют в кровеносные сосуды ножки гипофиза тиреолиберин, который, воздействуя на специфические клетки гипофиза, заставляет их высвобождать тиреотропный гормон (ТТГ). ТТГ током крови транспортируется к щитовидной железе и побуждает ее клетки синтезировать и выделять тиреоидный гормон. Этот гормон стимулирует разнообразные метаболические процессы в большинстве клеток организма. При этом повышение концентрации тиреоидного гормона в крови тормозит секрецию тиреолиберина и ТТГ (на схеме не показано). Эта отрицательная обратная связь препятствует чрезмерному повышению уровня тиреоидного гормона в крови. Подобного рода механизм регулирует секрецию многих гормонов.
|
|
|
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
| Таблица 12-1. Некоторые примеры внеклеточных сигнальных молекул | ||||
| Источник | Структура | Главные эффекты | ||
| Локальные химические медиаторы | ||||
| Белки | ||||
| Фактор роста нервов | Кожа; все ткани, иннервируемые симпатическими нервами | Две идентичные цепи из 118 аминокислот | Выживание и рост сенсорных и симпатических нейронов и некоторых нейронов в ЦНС | |
| Олигопептиды | ||||
| Фактор хемотаксиса эозинофилов | Тучные клетки | 4 аминокислоты | Хемотаксический сигнал для лейкоцитов определенного типа (эозинофилов) | |
| Производные аминокислот | ||||
| Гистамин | Тучные клетки | 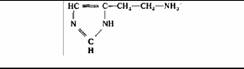
| Вызывает расширение кровеносных сосудов и повышение их проницаемости | |
| Производные жирных кислот | ||||
| Простагландин | Клетки многих типов | 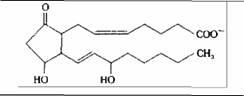
| Сокращение гладкой мускулатуры | |
| Нейромедиаторы * | ||||
| Аминокислоты и родственные соединения | ||||
| Глицин | Нервные окончания | 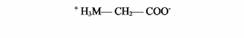
| Тормозный медиатор в ЦНС | |
| Норадреналин | Нервные окончания | 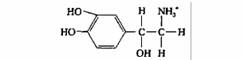
| Возбуждающий и тормозный медиатор центральной и периферической нервной системы | |
| γ-Аминомасляная кислота | Нервные окончания | 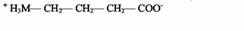
| Тормозный медиатор в ЦНС | |
| Ацетилхолин | Нервные окончания | 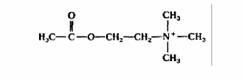
| Возбуждающий медиатор в нервно-мышечном соединении; возбуждающий и тормозный медиатор в центральной и периферической нервной системе | |
| Олигопептиды | ||||
| Энкефалин | Нервные окончания | 5 аминокислот | Подобно морфину, тормозит передачу болевых сигналов в ЦНС | |
| Гормоны2 | ||||
| Белки | ||||
| Инсулин | ß-Клетки поджелудочной железы | Белок: а-цепь-21 аминокислота, ß-цепь-30 аминокислот | Утилизация углеводов (включая захват глюкозы клетками); стимуляция белкового синтеза; стимуляция синтеза липидов в жировых клетках | |
| Соматотропин (гормон роста) | Передняя доля гипофиза | Белок, 191 аминокислота | Стимулирует синтез в печени соматомедина-1, вызывающего рост мышц и костей; сти- | |
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
| Продолжение табл. 12.1 | |||
| Источник | Структура | Главные эффекты | |
| мулирует дифференцировку жировой, мышечной и хрящевой ткани | |||
| Соматомедин- 1 (инсулино-подобный фактор роста 1) | Главным образом печень | Белок, 70 аминокислот | Рост костной и мышечной ткани; влияние на метаболизм Са2+, фосфата, углеводов и липидов |
| Адренокорти- котропный гормон (АКТГ) | Передняя доля гипофиза | Белок, 39 аминокислот | Стимуляция синтеза кортизола корой надпочечников; освобождение жирных кислот из жировых клеток |
| Паратгормон | Паращитовидные железы | Белок, 84 аминокислоты | Усиливает резорбцию кости, как следствие - повышает уровень Са2+ и фосфата в крови; усиливает реабсорбцию почками Са2+ и Mg2+ и уменьшает реабсорбцию фосфата |
| Фолликулостимулиру ющий гормон (ФСГ) | Передняя доля гипофиза | Гликопротеин, a-цепь-92 аминокислоты, ß-цепь 118 аминокислот | Стимуляция роста фолликулов яичника и секреции ими эстрадиола; стимуляция сперматогенеза в семенниках |
| Лютеинизирующий гормон (Л Г) | Передняя доля гипофиза | Гликопротеин, a-цепь-92 аминокислоты, ß-цепь - 115 аминокислот | Стимуляция созревания ооцитов, овуляции и секреции прогестерона яичником; стимуляция выработки тестостерона семенниками |
| Фактор роста эпидермиса | Неизвестен | Белок, 53 аминокислоты | Стимуляция клеток эпидермиса и других клеток к делению |
| Тиреотропный гормон (ттг) | Передняя доля гипофиза | Гликопротеин, a-цепь-92 аминокислоты, ß-цепь-112 аминокислот | Стимуляция синтеза тироксина в щитовидной железе; освобождение жирных кислот жировыми клетками |
| Олигопептиды | |||
| Тиреолиберин | Гипоталамус | 3 аминокислоты | Стимуляция синтеза тиреотропного гормона (ТТГ) передней долей гипофиза |
| Люлиберин | Гипоталамус | 10 аминокислот | Стимуляция синтеза лютеинизирующего гормона (ЛГ) передней долей гипофиза |
| Вазопрессин (антидиуретический гормон) | Задняя доля гипофиза | 9 аминокислот | Повышение кровяного давления в результате сужения мелких кровеносных сосудов; усиленная реабсорбция воды в почечных канальцах |
| Соматостатин | Гипоталамус | 14 аминокислот | Подавление секреции соматотропина передней долей гипофиза |
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
| Продолжение табл. 12.1 | |||
| Источник | Структура | Главные эффекты | |
| Производные аминокислот | |||
| Адреналин | Мозговое вещество надпочечников | 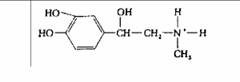
| Повышение кровяного давления, ускорение сердечного ритма; усиление гликогенолиза в печени и мышцах; выброс жирных кислот жировыми клетками |
| Тиреоидный гормон (тироксин) | Щитовидная железа | 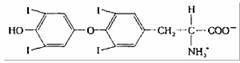
| Повышение метаболической активности большинства клеток |
| Стероидные гормоны | 
| ||
| Кортизол | Кора надпочечников | 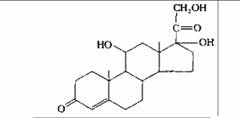
| Влияние на метаболизм белков, углеводов и липидов в большинстве тканей; подавление воспалительных реакций |
| Эстрадиол | Яичники, плацента | 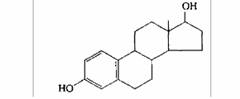
| Развитие и поддержание женских вторичных половых признаков; созревание и циклическая активность придаточных органов половой системы, развитие протоков молочной железы |
| Тестостерон | Семенники | 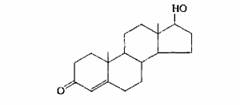
| Развитие и поддержание мужских вторичных половых признаков; созревание придаточных органов и поддержание их нормальной функции |
| Прогестерон | Яичники (желтое тело), плацента | 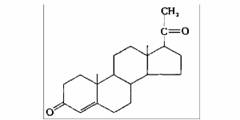
| Подготовка матки к беременности; сохранение беременности; развитие альвеолярной системы молочных желез |
| 1 Норадреналин и энкефалин чаще действуют по паракринному, чем по синаптическому типу; ацетилхолин может действовать и тем и другим способом. Возбуждающие нейромедиаторы стимулируют активность клеток-мишеней, а тормозные подавляют ее. | |||
| 2 Многие нестероидные гормоны синтезируются также некоторыми нейронами головного мозга. |
в общий кровоток.) Многие из гормонов гипофиза, выделяемых под контролем гипоталамуса, стимулируют какую-либо из других эндокринных желез и вызывают секрецию в кровь третьего гормона. Таким образом, гипоталамус служит у позвоночных главным регулятором эндокринной системы. В качестве примера на рис. 12-4 показано, каким образом он регулирует секреторную функцию щитовидной железы.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
В табл. 12-1 приведены сведения о некоторых локальных химических медиаторах, нейромедиаторах и гормонах - указаны места их синтеза, структура и основное действие. Видно, что структура сигнальных молекул так же разнообразна, как и их функции. Среди этих молекул есть короткие пептиды, более крупные белки и гликопротеины, аминокислоты и родственные им соединения, стероиды (вещества, образующиеся из холестерола и очень сходные между собой по структуре) и производные жирных кислот. Каждая сигнальная молекула представлена в табл. 12-1
только в одном разделе, хотя многие из них могут действовать несколькими способами. Например, некоторые пептидные гормоны в головном мозгу позвоночных действуют как нейромедиаторы (в качестве паракринных факторов).
Большинство клеток у взрослых животных специализировано для выполнения какой-то одной главной функции, и все они имеют характерный набор рецепторов, который позволяет им реагировать на химические сигналы, запускающие или модулирующие эту функцию. Многие сигнальные молекулы действуют в очень низкой концентрации (обычно не более 10-8 М), и связывающие их рецепторы, как правило, имеют к ним высокое сродство (константа сродства Ка ≥ 108 л/моль).
Одни и те же сигнальные молекулы часто оказывают различное действие на разные клетки-мишени. Например, ацетилхолин стимулирует сокращение волокон скелетной мускулатуры, но уменьшает частоту и силу сокращения клеток сердечной мышцы. В данном случае рецепторы ацетилхолина в скелетных мышцах отличаются от рецепторов на клетках миокарда. Но не всегда причина состоит в различии рецепторов. Часто одинаковые сигнальные молекулы связываются с идентичными рецепторами, и все же это ведет к совершенно разным реакциям у различных клеток-мишеней (рис. 12-5). Это означает, что ответы клеток-мишеней могут быть запрограммированы двумя способами: либо самим набором рецепторов клеточной поверхности, либо теми внутриклеточными системами, с которыми эти рецепторы сопряжены.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 1571; Нарушение авторских прав?; Мы поможем в написании вашей работы!