КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Актин и миозин создают силу, необходимую для цитокинеза
|
|
|
|
Митотическое веретено определяет место, где происходит разделение цитоплазмы при цитокинезе
13-30
Во время цитокинеза разделяется цитоплазма. Хотя деление ядра и разделение цитоплазмы, как правило, взаимосвязаны, эта связь не всегда неразрывна. Даже в норме за делением ядра может не следовать цитокинез. Например, в раннем зародыше Drosophila происходит В циклов ядерных делений без разделения цитоплазмы; в результате образуется одна большая клетка с 6000 ядер, расположенных в один слой около ее поверхности. Одноядерные клетки образуются позже при дроблении цитоплазмы вокруг всех этих ядер (разд. 16.5.2), Хотя митозу не всегда непосредственно сопутствует цитокинез, митотическое веретено играет важную роль в определении того, когда и как он будет происходить. Цитокинез обычно начинается в анафазе, продолжается во время телофазы и захватывает часть последующего

Рис. 13-65. Начало дробления яйца лягушки. Микрофотографии, полученные с помощью сканирующего электронного микроскопа.
Образование борозды обусловлено активностью сократимого кольца, находящегося под мембраной. А. Вид клеточной поверхности при малом увеличении. Б. Участок борозды при большом увеличении. (H.W. Beams. R.G. Kessel, Am. Sci. 64: 279-290, 1976.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
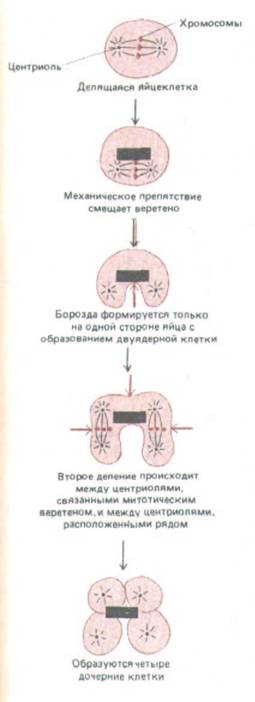
Рис. 13-66. Опыт, демонстрирующий влияние положения веретена на плоскость деления. Если митотическое веретено механически сместить на одну сторону клетки, то борозда дробления не дойдет до противоположной стороны клетки. Последующие деления будут происходить не только по экваторам двух митотических веретен (как это происходит в норме), но и между двумя соседними звездами, не связанными митотическим веретеном. Видимо, сократимый пучок из актиновых филаментов, создающий борозду дробления, всегда образуется в участке, лежащем посередине между двумя звездами. Это означает, что звезды каким-то образом изменяют окружающую область клеточного кортекса.
|
|
|
периода интерфазы. Первым видимым признаком цитокинеза у животных клеток бывает образование небольшой складки плазматической мембраны, появляющейся в анафазе и называемой бороздой деления (рис. 13-65). Эта борозда всегда образуется в плоскости метафазной пластинки, под прямым углом к длинной оси митотического веретена. Если в анафазе на достаточно раннем этапе веретено переместить с помощью микроманипулятора, то наметившаяся борозда исчезнет и появится новая в соответствии с новым положением веретена. Изящные опыты на яйцах морского ежа Echinarachnius показывают, что борозда дробления будет формироваться посередине между звездами, образовавшимися из двух центросом, даже если центросомы не связаны митотическим веретеном (рис. 13-66). Позднее, когда процесс зашел уже достаточно далеко, цитокинез будет продолжаться и в том случае, если веретено и его звезды удалить пипеткой или разрушить колхицином.
Большинство клеток делится симметрично. Борозда деления образуется по экватору родительской клетки, так что дочерние клетки будут одинаковой величины и с примерно одинаковыми свойствами. В период эмбрионального развития, однако, бывает много случаев, когда клетки делятся асимметрично: борозда разделяет две разные клетки, которые будут развиваться разными путями. Деления такого рода часто строго определены пространственно. Например, они могут происходить в определенных плоскостях по отношению к поверхности эпителиального пласта или приводить к обособлению участков цитоплазмы с разными наборами органелл. Независимо от того, будет ли деление симметричным или асимметричным, положение борозды, а значит, и плоскости деления всегда определяется положением митотического веретена. При надобности веретено может запрограммированным образом поворачиваться, занимая нужное положение в клетке и соответственно ориентируя плоскость деления (рис. 13-67). Кажется вероятным, что эти движения веретена определяются изменениями в отдельных участках клеточного кортекса, который сдвигает полюса веретена с помощью микротрубочек звезды. Видимо, сходный механизм определяет положение центросомы в поляризованной клетке (разд. 11.4.5). Структура кортекса, который богат актином, рассмотрена в гл. 11 (разд. 11.2).
|
|
|
Микротрубочки и пучки актиновых филаментов, находившиеся в интерфазной цитоплазме, во время митоза разрушаются. Однако цитоплазматические промежуточные филаменты во многих клетках сохраняются в целости. В таких клетках сеть промежуточных филаментов, окружающая интерфазное ядро, во время митоза растягивается, охватывая оба дочерних ядра, и в конце концов расчленяется на две части бороздой деления (рис. 13-68).
Разделение цитоплазмы происходит в результате сокращения кольца, состоящего главным образом из актиновых филаментов. Этот пучек
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
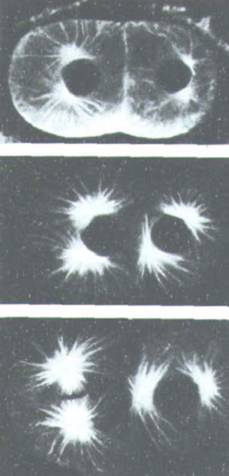
Рис. 13-67. Точно запрограммированный поворот митотического веретена на двуклеточной стадии зародыша нематоды Caenorhabditis
elegans при подготовке к делению с образованием четырех определенным образом расположенных клеток. (С любезного разрешения John White.)
филаментов, называемый сократимым кольцом (рис. 13-69), прикрепляется к внутренней стороне плазматической мембраны с помощью неидентифицированных белков. Сократимое кольцо образуется в начале анафазы, и механизм его сборки неизвестен; сила, которую оно создает, достаточна, чтобы согнуть тонкую стеклянную иглу, введенную в клетку. Нет сомнения в том, что источником силы здесь, так же как и в мышцах, служит взаимное скольжение актиновых и миозиновых филаментов. Например, в лизированных митотических клетках добавление субфрагментов инактивированного миозина блокирует миозин-связывающие участки актина, останавливая таким образом разделение цитоплазмы. Точно так же введение антител к миозину в яйца морского ежа вызывает сглаживание борозды дробления, но на ядерный митоз не влияет. Тем не менее в точности не известно, как взаимодействие актина и миозина втягивает плазматическую мембрану в борозду дробления.
|
|
|
В процессе нормального деления клетки сократимое кольцо не становится толще по мере углубления борозды. Это позволяет предполагать, что оно постепенно уменьшается в объеме за счет потери части филаментов. После завершения цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана в области борозды стягивается, окружая так называемое остаточное тельце, которое еще связывает две дочерние клетки. Остаточное тельце содержит остатки двух групп полярных микротрубочек, тесно упакованных вместе с материалом плотного матрикса (рис. 13-70).
Цитокинез, при котором из одной клетки образуются две, сильно увеличивает общую площадь клеточной поверхности. Поэтому двум дочерним клеткам требуется больше материала плазматической мембраны, чем исходной клетке. В животных клетках биосинтез вещества мембраны непосредственно перед делением усиливается. Избыточная мембрана у готовящихся к делению клеток, по-видимому, хранится в виде выступов (blebs) на их поверхности.
13.5.15. У высших растений цитокинез осуществляется совершенно иным способом [49]
Большинство клеток высших растений окружено жесткой клеточной стенкой, и поэтому механизм цитокинеза существенно отличается от только что описанного для животных клеток. Вместо образования двух дочерних клеток путем их отшнуровки с помощью сократимого кольца, лежащего под поверхностью клетки, цитоплазма разделяется здесь в результате образования новой стенки на границе между дочерними клетками.
|
|
|
Образующаяся перегородка точно определяет относительное положение двух новых клеток в растении. Из этого следует, что ориентация плоскостей клеточного деления и увеличение размеров клеток определяют форму растения (см. гл. 20).
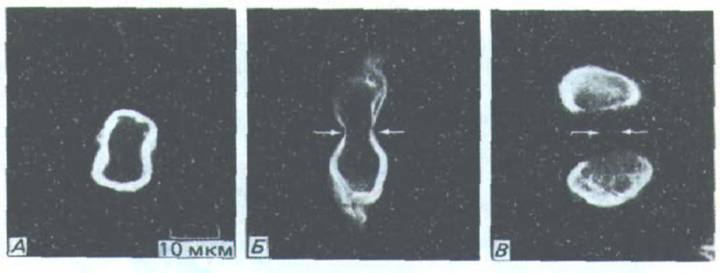
Рис. 13-68. Во время митоза зону ядра окружает пучок промежуточных филаментов. Микрофотографии получены после окрашивания пермеабилизиро ванных клеток флуоресцентными антителами, которые связываются с промежуточными филаменгами. А -анафаза; Б-ранняя телофаза {стрелками указано положение сократимого кольца); В-поздняя телофаза. (S. Н. Blose, Ргос. Natl. Acad. Sci. USA 76: 3372-3376, 1979.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
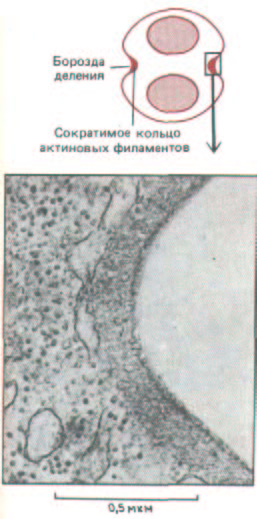
Рис. 13-69. Электронная микрофотография дна борозды, образующейся при делении животной клетки. Сверху для ясности приведена схема борозды деления. (Н. W. Beams, R. G. Kessel, Am. Sci. 64: 279-290, 1976.)
Новая поперечная перегородка, или клеточная пластинка, начинает строиться в плоскости между двумя дочерними ядрами в ассоциации с остаточными полюсными микротрубочками веретена, которые образуют цилиндрическую структуру, называемую фрагмопластом.
Эта структура, соответствующая микротрубочкам остаточного тельца животных клеток, состоит из двух групп противоположно ориентированных микротрубочек, расположенных параллельно друг другу (см. рис. 20-42). Микротрубочки, вероятно, прикреплены к поверхности ядра, так что их плюс-концы погружены в электроноплотный диск в экваториальной плоскости. Как показано на рис. 13-71, мелкие ограниченные мембраной пузырьки, происходящие в основном из аппарата Гольджи и наполненные предшественниками клеточной стенки, приходят в контакт с микротрубочками по обе стороны фрагмопласта и транспортируются вдоль них к экваториальной области клетки. Здесь они сливаются, образуя дисковидную, окруженную мембраной структуру - раннюю клеточную пластинку. Молекулы полисахаридов, высвобождаемые этими пузырьками, связываются между собой в ранней клеточной пластинке, образуя пектин, гемицеллюлозу и другие компоненты первичной клеточной стенки.
Теперь этот диск должен расширяться, пока его края не дойдут до стенки материнской клетки. Чтобы это стало возможным, микротрубочки раннего фрагмопласта претерпевают изменения по периферии ранней клеточной пластинки. Здесь с ними приходят в контакт новые пузырьки, которые затем сливаются на экваторе, расширяя пластинку. Этот процесс повторяется до тех пор, пока растущая клеточная пластинка не достигнет плазматической мембраны материнской клетки и мембраны не сольются, полностью разделяя две новые дочерние клетки (см. рис. 20-41 и 20-42).
Затем в клеточной пластинке
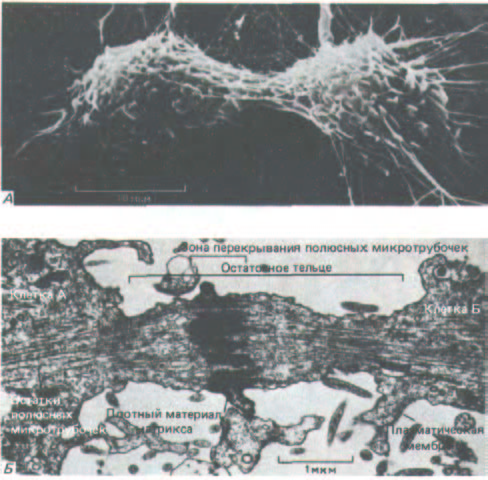
Рис. 13-70. А, Животная клетка в культуре в конце: деления: остаточное тельце остается связанным с обеими дочерними клетками.
Микрофотография, полученная с помощью сканирующего электронного микроскопа. Б. Электронная микрофотография остаточного тельца делящейся животной клетки. Деление практически полностью завершено, но дочерние клетки еще связаны тонким цитоплазматическим мостиком.
[С любезного разрешения Guenter Albrecht-Buehler (A) и J.M. Millins (Б).]
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
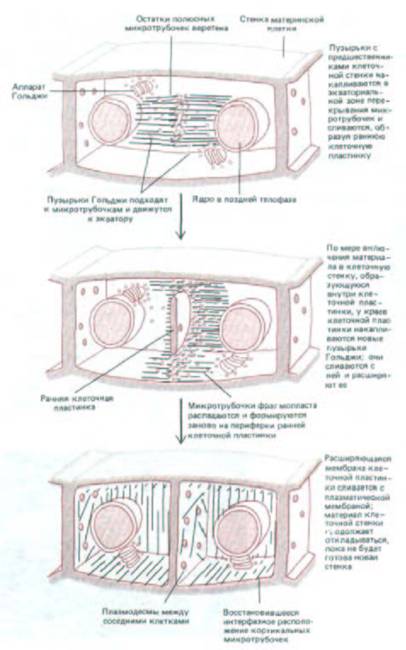
Рис. 13-71. Ход цитокинеза в клетках высших растений, имеющих жесткую клеточную стенку.
откладываются микрофибриллы целлюлозы, завершая построение новой клеточной стенки (рис. 13-71 и 13-72).
С пузырьками образующейся клеточной пластинки связаны элементы эндоплазматического ретикулума, которые часто оказываются включенными в эту пластинку. Впоследствии они превращаются в плазмодесмы - сложно устроенные поры, пронизывающие зрелую клеточную
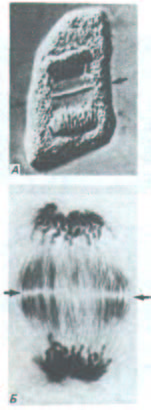
Рис. 13-72. Цитокинез в растительной клетке. Клеточная пластинка (между двумя стрелками) формируется в плоскости, перпендикулярной плоскости рисунка. Одна клетка (А) сфотографирована с применением метода дифференциального интерференционного контраста; другая окрашена антителами со связанными частицами золота, которые метят два скопления микротрубочек, входящих в состав фрагмопласта. В обоих случаях стрелками указана плоскость клеточной пластинки. [С любезного разрешения Jeremy D. Pickett-Heaps (А) и Andrew Bajer (Б).]
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 13-73. Организация актиновых филаментов в растительной клетке во время цитокинеза. Актиновые филаменты (выделенные темно-
красным цветом) формируют радиальную сеть, которая простирается от концов фрагмопласта до клеточного кортекса, образуя вокруг клетки кольцо. Эта сеть, по-видимому, определяет плоскость образования клеточной пластинки. Другая группа актиновых филаментов расположена параллельно микротрубочкам, участвующим в образовании новой клеточной пластинки в фрагмопласте. Еще одна группа актиновых филаментов (на рисунке не показана) подходит к кортексу из области двух дочерних ядер через большую центральную вакуоль, свойственную растительным клеткам (разд. 20.40.7); эти филаменты помогают поддерживать тонкие цитоплазматические мостики, пересекающие вакуоль.
стенку и соединяющие цитоплазму всех клеток растения (см. разд. 20.2.1 и рис. 20-20).
Так же как и у животных, митоз и цитокинез у растений могут быть разобщены. Так, например, в эндосперме семян митозы происходят без цитокинеза, что приводит к образованию гигантской многоядерной клетки. Значительно позднее, когда митотическое веретено уже давно распалось, между отдельными ядрами строятся новые клеточные стенки, так что образуются отдельные клетки.
Для определения точного положения и формы клеточной стенки одного митотического веретена обычно не достаточно. Место соединения будущей пластинки со стенкой материнской клетки, по-видимому, определяется очень рано, еще до начала митоза, узким пучком микротрубочек - предпрофазным пояском, расположенным непосредственно под плазматической мембраной (см. разд. 20.5.5 и рис. 20-64). Хотя эти микротрубочки в начале митоза исчезают, от них зависит, в каком участке кортекса будет прикрепляться радиальная сеть актиновых филаментов, которая сохраняется на протяжении всей фазы М и должна будет направлять растущий край клеточной пластинки к надлежащей зоне кортекса (рис.
13-73). Таким образом, актин играет важную роль и в делении клеток с жесткими стенками, где активное сокращение не играет, по-видимому, никакой роли. Поскольку актин участвует также в формировании клеточных септ у грибов, возможно, что он направляет цитокинез у всех эукариот.
13.5.16. Цитокинез должен обеспечить правильное распределение цитоплазматических органелл [50]
Ядро-это только одна из многих клеточных органелл, для удвоения которых необходима предшествующая органелла того же типа.
Например, рибосомы могут спонтанно собираться из своих компонентов, но для их построения нужны другие рибосомы, чтобы синтезировать необходимые белки. С другой стороны, митохондрии и хлоропласта не способны к спонтанной самосборке и могут образовываться только путем роста и разделения предсуществующих органелл (разд. 7.5.1). Точно так же механизмы роста ряда других органелл, например аппарата Гольджи и эндоплазматического ретикулума, таковы, что трудно представить себе их новообразование в отсутствие хотя бы фрагментов соответствующих структур (см. гл. 8). У некоторых водорослей, имею-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
щих только один хлоропласт или только один аппарат Гольджи, органелла, присутствующая в одном-единственном экземпляре, перед цитокинезом расщепляется на две половинки, которые затем и распределяются между дочерними клетками (см. рис. 7-67). Примером такого же явления служат дупликация и сегрегация центросомы в животных клетках (см. рис. 13-46).
Как же при делении клеток высших эукариот разделяются различные органеллы, окруженные мембраной (за исключением ядра)? В большинстве случаев число этих органелл достаточно велико (см. табл. 8-1), чтобы и при случайном распределении их в процессе цитокинеза каждая дочерняя клетка получала их более или менее представительный набор. Таким образом, хотя клетка млекопитающего не выживет, не получив, например, ни одной митохондрии, вполне возможно, что для надежной передачи их дочерним клеткам не требуется никакого специального механизма. Разумеется, органеллы, присутствующие в клетках в большом количестве, будут всегда успешно наследоваться, если в среднем их число будет удваиваться в каждом клеточном поколении. Другие органеллы, такие как аппарат Гольджи и эндоплазматический ретикулум, во время митоза распадаются на более мелкие фрагменты и пузырьки. Такое раздробление, вероятно, способствует их равному распределению между дочерними клетками.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 1020; Нарушение авторских прав?; Мы поможем в написании вашей работы!