КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Прикрепительные контакты связывают цитоскелет клетки с цитоскелетом соседней клетки или с
|
|
|
|
14-5
14-4
14-8
14.1. Межклеточные соединения [1]
Специализированные межклеточные соединения особенно многочисленны и важны в эпителиях, но во многих местах контакта между клетками и между клетками и матриксом они встречаются во всех тканях. В большинстве своем они слишком малы для того, чтобы их можно было увидеть в световой микроскоп; однако их можно выявить с помощью электронной микроскопии в обычных препаратах или же в препаратах, полученных методом замораживания-скалывания. В обоих случаях видно, что взаимодействующие плазматические мембраны (а нередко и подстилающие их участки цитоплазмы и межклеточное пространство) имеют в этих местах высокоспециализированную структуру. Клеточные соединения могут быть разделены на три функциональные группы: 1) запирающие соединения, которые так тесно сцепляют клетки в эпителиальном пласте, что делают невозможным прохождение даже небольших молекул с одной стороны пласта на другую; 2) прикрепительные
контакты, которые механически связывают клетки (и их цитоскелеты) с соседними клетками или внеклеточным матриксом; и 3)
коммуникационные контакты, по которым передаются химические или электрические сигналы между взаимодействующими клетками.
Основные типы межклеточных соединений внутри каждой из этих групп перечислены в табл. 14-1. Основным типом замыкающих контактов являются плотные соединения; адгезионные соединения и десмосомы составляют основной тип прикрепительных контактов; а щелевые
кон-
Таблица 14-1. Функциональная классификация клеточных соединений __________________________________________________________________
I. Замыкающие (плотные) соединения
II. Прикрепительные соединения
|
|
|
1. Места прикрепления актиновых филаментов (адгезионные контакты)
а) между клетками (например, адгезионные пояса)
б) между клетками и матриксом (например, фокальные контакты)
2. Места прикрепления промежуточных филаментов а) между клетками (десмосомы)
б) между клетками и матриксом (полудесмосомы)
III. Коммуникационные соединения
1. Щелевые контакты 2. Химические синапсы 3. Плазмодесмы (только у растений)1)
___________
1) Это единственный род соединений между растительными клетками.
__________________________________________________________________
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
такты, химические синапсы нервных клеток и, наконец, плазмоде с мы, образующиеся между растительными клетками, - это главные виды коммуникационных контактов. Так как химические синапсы и плазмодесми будут детально рассматриваться в гл. 19 и 20 соответственно, мы не будем обсуждать их в этой главе.
14.1.1. Плотные соединения создают в эпителиальных клеточных пластах барьер проницаемости [2]
Несмотря на существенные структурные и биохимические различия между разными типами эпителиев, эти ткани обладают по меньшей мере одной общей функцией: они служат барьерами с избирательной проницаемостью, разделяющими жидкости разного химического состава по обе стороны пласта. Плотные соединения играют вдвойне важную роль в поддержании такой барьерной функции. Это хорошо иллюстрируется на примере кишечного эпителия млекопитающих.
Эпителиальные клетки, выстилающие тонкий кишечник, удерживают большую часть содержимого кишки в ее внутренней полости (просвете). Однако в то же время они должны перекачивать определенные питательные вещества через клеточный пласт во внеклеточную жидкость соединительной ткани по другую сторону пласта (см. рис. 14-1), откуда эти вещества диффундируют в кровеносные сосуды. Такой трансэпителиальный перенос осуществляют две группы транспортных белков, связанных с мембраной. Одна группа расположена на апикальной
|
|
|
поверх-
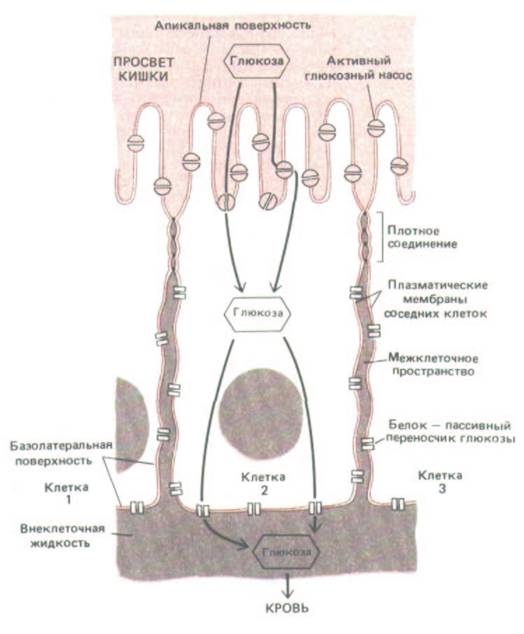
Рис. 14-2. Схема эпителиальной клетки из тонкой кишки: показано, как плотные контакты разграничивают области плазматической мембраны, в которых могут находиться различные транспортные белки. Такое разграничение обеспечивает перенос питательных веществ из просвета кишки через эпителиальный слой в кровь. В представленном здесь примере глюкоза активно транспортируется в клетку глюкозними насосами апикальной поверхности, а затем выходит из клетки путем облегченной диффузии при участии белков - пассивных переносчиков глюкозы, находящихся в базолатеральной области мембраны. Плотные соединения, по-видимому, ограничивают перемещение белков определенными участками плазматической мембраны, действуя как диффузионные барьеры внутри ее липидного бислоя; эти соединения блокируют также диффузию липидных молекул в наружном (но не во внутренном) листке липидного бислоя.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 14-3. Растворимая меченая молекула, введенная по одну сторону эпителиального слоя, не может пройти через плотные соединения, скрепляющие соседние клетки. Но это препятствие не абсолютно, и есть данные о том, что клетки могут изменять свойства своих плотных соединений для регулирования потока растворенных веществ и воды через эпителий.
ности (обращенной в просвет кишки) и активно переносит избранные молекулы из просвета в эпителиальные клетки; другая группа расположена на базолатеральных (базальных и латеральных) поверхностях клеток и позволяет тем же молекулам выходить из клеток, облегчая диффузию во внеклеточную жидкость по другую сторону эпителия (рис. 14-2). Для поддержания такого направленного транспорта нужно, чтобы апикальные транспортные белки не могли переходить на базолатеральную поверхность клетки, а базолатеральные - на апикальную поверхность. Кроме того, щели между эпителиальными клетками должны быть так закупорены, чтобы транспортируемые молекулы не могли диффундировать обратно в полость кишки через межклеточные пространства «вниз» по градиентам концентрации, создающимся в результате трансэпителиального переноса.
|
|
|
По-видимому, плотные соединения между эпителиальными клетками препятствуют обоим этим видам диффузии. Во-первых, они действуют как барьеры для диффузии мембранных белков между апикальной и базолатеральной поверхностями плазматической мембраны (рис.
14-2). Такая нежелательная диффузия компонентов мембраны происходит при разрушении плотных соединений, например при удалении внеклеточных ионов Са2 +, необходимых для сохранения целостности плотного соединения. Во-вторых, соседние клетки оказываются так плотно сомкнутыми, что даже и водорастворимые молекулы не прохо-

Рис. 14-4. Структура плотного соединения между эпителиальными клетками тонкой кишки. А, Схема. Б. Электронная микрофотография препарата, полученного методом замораживания-скалывания. В. Обычная электронная микрофотография. Обратите внимание, что клетки ориентированы апикальными концами вниз. На фото Б плоскость микрофотографии параллельна плоскости мембраны; видно, что плотное соединение образовано сетью из герметизирующих цепочек, опоясывающей каждую клетку в пласте. Эти герметизирующие цепочки видны как гребни из внутримембранных частиц на внутренней (цитоплазматической) поверхности скола (В) или как комплементарные им бороздки на наружной поверхности мембраны (Н). На обычном препарате (В) соединение выглядит как серия фокальных контактов между наружными липидными слоями двух смежных мембран; каждый такой контакт соответствует герметизирующей цепочке в поперечном разрезе. [Б и В из N. В.
Giluda. Tn: Cell Communication (R.P, Cox, ed.), pp. 1-29. New York, Wiley, 1974. Reprinted by permission of John Wiley a. Sons, Inc.]
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
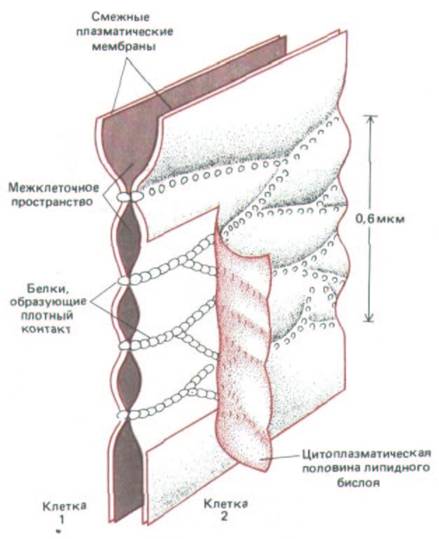
Рис. 14-5. Современная модель строения плотного соединения. Предполагается, что смежные плазматические мембраны скреплены непрерывными цепочками из особых трансмембранных белков, осуществляющих контакт через межклеточное пространство и образующих герметичное соединение. Чтобы показать эти белковые цепочки, внутренний липидный монослой одной из мембран на этой схеме отогнут. На препаратах, приготовленных методом замораживания -скалывания, белки плотного соединения остаются не на наружном монослое мембраны, как показано здесь, а на внутреннем, где создают узор внутримебранных частиц, показанный на рис. 14-4, Б.
дят между ними: если с одной сторони эпителиального клеточного пласта ввести электроноплотный маркер из малых молекул, то обычно он не проникает через плотное соединение (рис. 14-3).
Молекулярная структура плотного соединения еще не ясна, но электронная микроскопия с применением метода замораживания -
скалывания показывает, что оно состоит из сети анастомозирующих волокон, которая оплетает апикальный конец каждой клетки по всей его окружности (рис. 14-4, А и Б). На обычных электронных микрофотографиях они видны как серии локальных соединений между наружными поверхностями двух смежных плазматических мембран (рис. 14-4, В). Хотя все плотные соединения непроницаемы для макромолекул, их проницаемость для малых молекул сильно различается у разных эпителиев. Например, в эпителии, выстилающем тонкий кишечник, плотные соединения в 10000 раз более проницаемы для ионов, чем в эпителии мочевого пузыря. Способность соединения препятствовать переходу ионов через межклеточные пространства увеличивается в логарифмической зависимости от числа волокон в сети, как если бы каждое волокно действовало как независимый барьер. Как полагают, волокна состоят из длинных рядов специфических трансмембранных белков каждой из двух контактирующих мембран, которые (белки) непосредственно соединяются друг с другом, замыкая межклеточное пространство (рис. 14-5).
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2871; Нарушение авторских прав?; Мы поможем в написании вашей работы!