КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Внеклеточный матрикс состоит в основном из фибриллярных белков, погруженных в гидратированный
|
|
|
|
14-7
14.1.7. Большинство клеток в ранних эмбрионах сообщается через щелевые контакты [7]
В некоторых тканях роль сопряжения клеток через щелевые контакты очевидна. Например, электрическое сопряжение синхронизирует сокращения клеток сердечной мышцы и клеток гладкой мускулатуры, ответственных за перистальтику кишечника. Точно так же электрическое сопряжение между нервными клетками позволяет потенциалам действия быстро распространяться от клетки к клетке без задержки, происходящей в химических синапсах; это дает преимущество в случаях, когда решающее значение имеют быстрота и надежность ответа, например при некоторых реакциях бегства у рыб и насекомых. Труднее понять, зачем нужны щелевые контакты в тканях, не проявляющих электрической активности. В принципе обмен метаболитами и ионами мог бы обеспечить координацию активности отдельных клеток в этих тканях. Например, через щелевые контакты могла бы координироваться такая активность клеток эпителиального слоя, как биение ресничек; а поскольку внутриклеточные посредники типа циклического AMP способны проходить через щелевые контакты, ответ сопряженных клеток на внеклеточные сигнальные молекулы мог бы распространяться и координироваться именно этим путем.
По-видимому, сопряжение клеток через щелевые контакты играет важную роль в эмбриогенезе. В ранних зародышах позвоночных (у мышиного - начиная с поздней стадии восьми бластомеров) большинство клеток электрически связано друг с другом. Однако по мере того, "как специфические группы клеток приобретают явные различия и начинают дифференцироваться, они обычно утрачивают сопряжение с окружающими тканями. Например, при замыкании нервной трубки ее клетки теряют связь с покрывающей эктодермой (см. рис. 14-9). Тем временем клетки внутри каждой группы остаются сопряженными друг с другом и поэтому ведут себя как кооперативная система, согласованно следуя по определенному пути развития.
|
|
|
Одна из привлекательных гипотез состоит в том, что сопряжение эмбриональных клеток могло бы обеспечивать возможность дальнодействуюшей сигнализации в развивающемся эпителии. Например, малые молекулы могли бы переходить через щелевые контакты из тех участков ткани, где их концентрация поддерживается на высоком уровне, в участки, где она остается низкой, так что создавался бы плавный градиент. Локальный уровень концентрации мог бы доставлять клеткам «позиционную информацию» для управления их дифференцировкой в соответствии с их локализацией в зародыше. Но действительно ли щелевые контакты выполняют такую функцию, не известно.
На возможную роль межклеточной коммуникации через щелевые контакты в процессах развития указывают эксперименты, в которых в один из бластомеров 8-клеточного зародыша амфибии инъецировали антитела к главному белку щелевого контакта. Введенные антитела не только избирательно разрывали электрическое сопряжение и предотвращали перенос красителя между потомками обработанной клетки (что проверялось через два цикла деления - на 32-клеточной стадии), но и резко нарушали развитие зародыша (рис. 14-18). Остается неясным, каким образом разрыв клеточного сопряжения на ранней стадии приводит позже к дефектам в развитии, однако эксперименты такого рода - многообещающий первый шаг в изучении роли щелевых контактов в эмбриональном развитии.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
14.1.8. Проницаемость щелевых контактов может регулироваться [8]
Экспериментальные воздействия, снижающие рН или повышающие концентрацию свободных ионов Са2 + в цитозоле, быстро (за несколько секунд) и обратимо уменьшают проницаемость щелевых контактов, а в некоторых тканях проницаемость их может регулироваться градиентом напряжения на контакте или внеклеточными химическими сигналами. Эти наблюдения показывают, что щелевые контакты -
динамичные структуры, способные открываться или закрываться в ответ на изменения в клетках. Таким образом, в этом отношении они сходны с обычными ионными каналами (разд. 6.4.14), хотя переходы между открытым и закрытым состояниями происходят здесь значительно реже, чем у большинства ионных каналов.
Какую роль играет регуляция проницаемости щелевых контактов потенциалом или величиной рН в нормальном функционировании клеточных ансамблей, не известно. В одном случае однако, смысл контроля с участием ионов Са2+ кажется понятным. При гибели или повреждении клетки ее мембрана утрачивает барьерную функцию. Такие ионы, как Са2+ или Na +, входят в клетку, а важные метаболиты выходят из нее. Если бы такая клетка оставалась связанной со своими здоровыми соседями, то их внутренняя среда тоже подвергалась бы опасности. Однако повышение концентрации Са2+ в поврежденной клетке приводит к закрытию каналов щелевых контактов, что эффективно изолирует ее и таким образом предотвращает распространение повреждения.
Повышение проницаемости щелевых контактов под действием внеклеточных химических сигналов ведет к распространению реакции на соседние клетки, не находящиеся в прямом контакте с действующим агентом. Например, гормон глюкагон, побуждающий клетки печени к расщеплению гликогена и высвобождению глюкозы в кровяное русло, может также повышать проницаемость щелевых контактов между этими клетками у крысы. Это происходит за счет увеличения внутриклеточной концентрации циклического AMP (сAMP), который активирует сАМР-
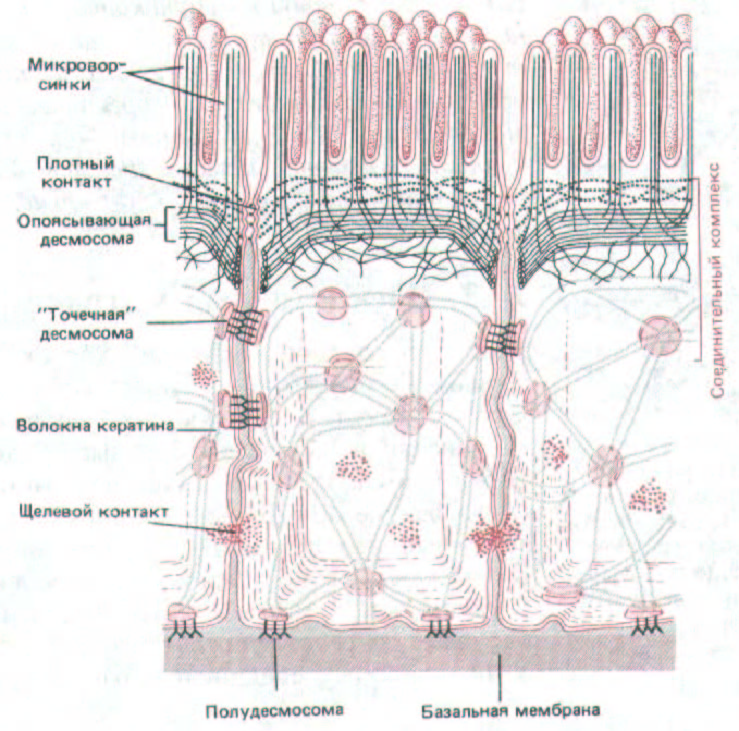
Рис. 14-19. Схема расположения различных соединений, образуемых эпителиальными клетками тонкой кишки.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
зависимую протеинкиназу (разд. 12.4.1), а та в свою очередь, по-видимому, фосфорилирует главный белок щелевого контакта. Расщепление гликогена клетками печени тоже обусловлено повышением концентрации сАМР, так что одновременное увеличение проницаемости щелевых контактов, облегчая диффузию сАМР из клетки в клетку, способствует вовлечению соседних групп клеток в процесс расщепления гликогена. На рис. 14-19 суммированы различные типы соединений, образующихся между клетками в эпителии. В апикальном конце клетки относительное положение клеточных соединений одинаково почти во всех эпителиях: плотные соединения занимают наиболее апикальную область клетки, за ними идет адгезионный пояс, а дальше - специальные параллельные ряды десмосом; все вместе они образуют «соединительный комплекс». Менее регулярно располагаются щелевые контакты и дополнительные десмосомы.
Заключение
Большинство клеток в тканях связаны друг с другом и с внеклеточным матриксом в специализированных местах контакта, называемых клеточными соединениями. Клеточные соединения разделяют на три функциональных класса: запирающие, прикрепительные и
коммуникационные. Плотные соединения составляют главную группу запирающих соединений и играют основную роль в поддержании разности
концентраций малых гидрофильных молекул по разные стороны эпителиальных слоев; они, во-первых, плотно связывают мембраны соседних
клеток и создают таким образом непрерывный барьер проницаемости между двумя сторонами эпителия и, во-вторых, образуют барьер в
липидном бислое, предотвращающий диффузию мембранных транспортных белков между апикальной и базо латеральной областями
плазматической мембраны каждой эпителиальной клетки.
Существуют два основных типа прикрепительных контактов: адгезионные соединения и десмосомы. Все они объединяют группы
клеток в прочные структурные комплексы, связывая элементы их цитоскелетов. Адгезионные соединения связывают пучки актиновых
|
|
|
филаментов, а десмосомы-промежуточные филаменты. Щелевые контакты служат для межклеточной коммуникации и состоят из групп
канальных белков, позволяющих частицам с мол. массой менее 1500 непосредственно переходить из одной клетки в другую. Клетки, связанные
такими контактами, обмениваются многими неорганическими ионами и другими малыми молекулами, т, е. они химически и электрически
сопряжены. Щелевые контакты имеют большое значение для координации функций электрически активных клеток и, по-видимому, играют
сходную роль также в других группах клеток.
14.2. Внеклеточный матрикс [9]
Ткани состоят не только из клеток. Значительную часть их объема занимает внеклеточное пространство, заполненное сложной сетью макромолекул, составляющих внеклеточный матрикс (рис. 14-20). Этот матрикс включает разнообразные полисахариды и белки, которые секретируются самими клетками и организуются в упорядоченную сеть. Описывая межклеточные соединения, мы рассматривали главным образом эпигелиальные ткани, при описании же внеклеточного матрикса мы будем иметь дело в основном с соединительными тканями (рис. 14-21). В таких тканях матрикс обычно занимает больший объем, чем клетки, окружает их со всех сторон и определяет механические свойства ткани. У позвоночных соединительные ткани образуют структурный каркас
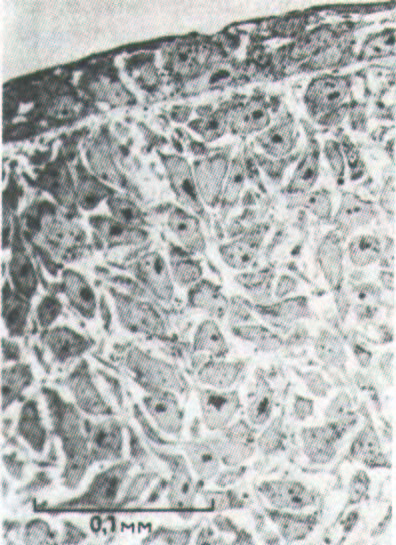
Рис. 14-20. Электронная микрофотография, показывающая при небольшом увеличении клетки, окруженные внеклеточным матриксом. В данном случае это еще не дифференцированные клетки конечности раннего куриного эмбриона. (С любезного разрешения Cheryll Tickle.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
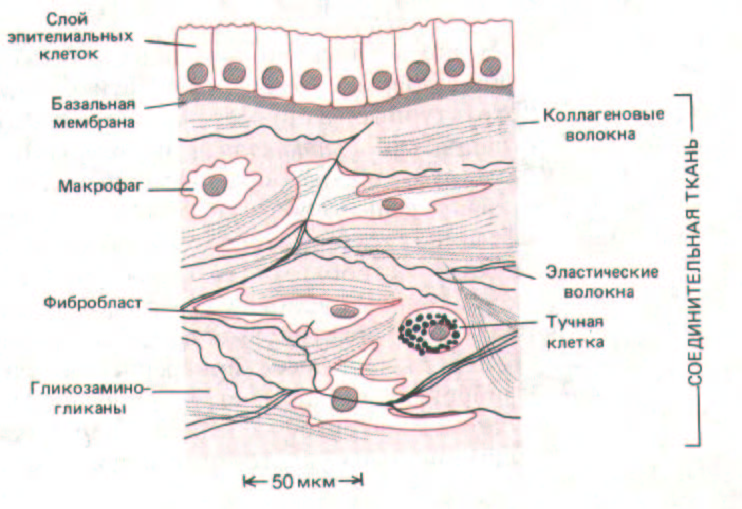
Рис. 14-21. Соединительная ткань, подстилающая слой эпителиальных клеток.
тела, но их доля в разных органах весьма различна: в коже и костях, например, они являются основным компонентом, а в головном и спинном мозгу составляют лишь незначительную часть.
Различия в соотношениях разных типов макромолекул и в способе их организации во внеклеточном матриксе порождают необычайное разнообразие форм, каждая из которых очень хорошо приспособлена к функциональным потребностям данной ткани. Матрикс может обызвествляться, образуя твердые, как камень, структуры кости или зуба, может формировать прозрачное вещество роговицы глаза или принимать форму каната, что придает сухожилиям огромную прочность на разрыв. На границе между эпителием и соединительной тканью матрикс образует базальную мембрану - чрезвычайно тонкую, но плотную прокладку, играющую важную роль в регуляции поведения клеток. Мы ограничимся описанием внеклеточного матрикса позвоночных, но интересные и своеобразные структуры того же рода встречаются и у многих других организмов; таковы, например, клеточные стенки бактерий и растений, кутикула червей и насекомых, раковины моллюсков. Стенки растительных клеток будут детально рассмотрены в гл. 20.
До недавнего времени внеклеточный матрикс позвоночных считали сравнительно инертным каркасом, стабилизирующим физическую структуру тканей. Но сейчас стало ясно, что он играет значительно более активную и сложную роль в регуляции поведения контактирующих с ним клеток - влияет на их развитие, миграцию, пролиферацию, форму и метаболизм. Молекулярный состав внеклеточного матрикса достаточно сложен, но, хотя наше понимание его организации пока еще фрагментарно, идет быстрый прогресс в изучении его главных компонентов.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 814; Нарушение авторских прав?; Мы поможем в написании вашей работы!