КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Основи молекулярної біології та молекулярної генетики
|
|
|
|
РОЗДІЛ 2
Основи молекулярної біології та молекулярної генетики
РОЗДІЛ 2.
2.1.Будова та властивості молекулиДНК
2.2.Передача генетичної інформації. Мутації
2.3.Розшифрування генетичної інформації
2.3.1. Транскрипції
2.3.2.Трансляція
2.4.Технологія рекомбінантних ДНК
2.4.1.Ферменти генної інженерії
2.4.2.Будова рестрикційних карт
2.4.3.Визначення нуклеотидної послідовності ДНК
2.4.4.Методи конструювання рекомбінантних ДНК
2.4.5.Векторні молекули
2.4.6.Введення молекул ДНК у клітини
2.4.7.Створення і скринінг геномних бібліотек
Контрольні запитання
Живий організм являє собою відкриту термодинамічну систему, що само відтворюється, у якій шляхи перетворення речовини й енергії обумовлюються генетичною інформацією, що реалізується через генетичний код. У такому визначенні М. Барб'єрі (1998 р.) сформулював один з фундаментальних принципів біології кінця XXст. Можливість появи цього глобального узагальнення пов'язана з трьома найбільшими досягненнями в біологічних дослідженнях попереднього століття. Біохіміками було визначено функціональну роль білків, генетикам удалося підняти завісу над світом генетичної інформації, а молекулярні біологи встановили, що зв'язок між цими двома світами здійснюється через генетичний код.
Відповідно до сучасних уявлень уся генетична інформація живого організму утримується в його генах і, як будь-яка інша інформація, містить у собі повідомлення, у цьому разі, повідомлення для молекулярних об'єктів, здатних їх сприйняти. Сама можливість сприйняття генетичної інформації визначається тим, що повідомлення організовані за допомогою системи правил (генетичного коду), «зрозумілих» об'єктам, для яких ці повідомлення призначені. Отримавши генетичне повідомлення, молекулярний об'єкт його декодує відповідно до правил, що лежать в основі функціонування його складових частин. Навіть часткове невиконання цих правил може призвести до важких порушень життєдіяльності організму.
|
|
|
Під час передачі генетичної інформації з існуючих каналів зв'язку від генів до сприймаючих молекулярних об'єктів має місце її багаторазове декодування й перекодування аж до остаточного втілення у фенотипічних ознаках. Відбувається експресія генів, тобто реалізація генетичної інформації.
Клітина є тією найменшою частиною організованої матерії, що ще зберігає всі ознаки живого. У ній відбуваються основні події, пов'язані з експресією генів. Лише в клітині може повноцінно реалізовуватися генетична інформація, що є в генах. Зв'язок між генами й внутрішньоклітинним генетичним оточенням нерозривний. Компоненти клітини, розпізнаючи гени, зчитують укладену в них інформацію й декодують її. При цьому вони підтримують гени в робочому стані і їх відтворюють, створюючи точну копію носія інформації, гарантію існування самих себе.
«Omniameamecumporto!» ( Все своє ношу із собою лат.) цей девіз повною мірою стосується одноклітинних організмів. У багатоклітинних генетичних систем це не так. У цьому разі окремі клітини, що складають організм, стають залежними один від іншого, створюючи нерозривну єдність на певному рівні − соми, або тіла. Між клітинами відбувається контрольований генами перерозподіл функцій. При цьому клітини різних частин багатоклітинного організму можуть настільки розрізнятися морфологічно, що без проведення спеціального аналізу на молекулярному рівні їх неможливо віднести до однієї генетичної системи, єдиного організму.
Перехід до багатоклітинності змінив стосунки організму з навколишнім світом на новий рівень складності. Численні зв'язки між рецепторами організму й зовнішніми впливами, з одного боку, а також внутрішніми аналізаторами інформації, що надходить, з іншого забезпечують максимальну пристосованість організму до умов існування. Життя не прощає помилок. Усе, що неадекватно реагує на сигнали середовища існування, елімінується (виключається, видаляється) природнім відбором. Чим ефективніше система налаштована на навколишній світ (у тому числі й на внутрішнє середовище організму), тим складніше й витонченіше її морфологічне втілення, її внутрішній зміст.
|
|
|
В основі розвитку будь-якого багатоклітинного організму, становлення системи його органів і тканин лежить поділ (проліферація) клітин. Генетична програма забезпечує протікання складної сукупності біохімічних реакцій, що супроводжують створення точної копії генетичного апарату кожної соматичної клітини, її ріст і подвоєння. Оскільки за кожного ділення клітин весь глобальний біохімічний процес циклічно повторюється, він дістав назву клітинного циклу. Індивідуальний (онтогенетичний) розвиток, як правило, починається з першого подвоєння стимульованого до цього яйця (яйцеклітини) і завершується тільки з настанням його смерті − розпаду організму як цілого внаслідок обриву ключових внутрішніх зв'язків між системами його життєзабезпечення. Основна внутрішня подія життя
організму − поділ клітин − є під суворим внутрішнім і зовнішнім генетичним контролем. Навіть ізольована соматична клітина здатна лише до обмеженої кількості поділів у поживному середовищі. Кількісний контроль числа клітинних поділів лежить в основі органогенезу (формування органів і тканин). Порушення механізмів контролю проліферації клітин призводить до нестримного подвоєння клітин, утворення безформної клітинної маси − пухлини, здатної задушити організм зсередини. Однак, у процесі нормального онтогенетичного розвитку змінюється не тільки число соматичних клітин, а й їх якісний склад.
Здатність органів і тканин здійснювати свої специфічні функції цілком залежить від наявності в них спеціалізованих клітин. Зокрема,організм дорослої людини складається з  1014…1015 клітин більш ніж 100 різних типів. На дуже ранніх стадіях розвитку зародка багатоклітинного організму клітини, з яких він складається, зовні схожі одна на одну. У міру продовження онтогенезу шляхи багатьох з них далеко розходяться. Відбувається диференціація клітин, набуття ними спеціалізованих функцій. Морфологічні різниці, що виявляють у спеціалізованих клітин, обумовлюються особливим складом і внутрішньоклітинною організацією їх молекул. Поява таких особливостей на молекулярному рівні також контролюється генами. У спеціалізованих (диференційованих) клітинах або їхніх попередниках крім генів, що здатні до експресії в клітинах усіх типів, працюють особливі групи генів. Перехід експресії одних груп генів на інші, залучення до експресії нових генів і припинення роботи старих у клітинах, що диференціюються, також перебуває під точним генетичним контролем.
1014…1015 клітин більш ніж 100 різних типів. На дуже ранніх стадіях розвитку зародка багатоклітинного організму клітини, з яких він складається, зовні схожі одна на одну. У міру продовження онтогенезу шляхи багатьох з них далеко розходяться. Відбувається диференціація клітин, набуття ними спеціалізованих функцій. Морфологічні різниці, що виявляють у спеціалізованих клітин, обумовлюються особливим складом і внутрішньоклітинною організацією їх молекул. Поява таких особливостей на молекулярному рівні також контролюється генами. У спеціалізованих (диференційованих) клітинах або їхніх попередниках крім генів, що здатні до експресії в клітинах усіх типів, працюють особливі групи генів. Перехід експресії одних груп генів на інші, залучення до експресії нових генів і припинення роботи старих у клітинах, що диференціюються, також перебуває під точним генетичним контролем.
|
|
|
Ще однією важливою подією індивідуального розвитку організму є повне заміщення одних груп клітин іншими. При цьому кінцеві стадії процесу заміни контролюються самими клітинами, що заміщуються. На певній стадії розвитку ембріона усередині клітин, які видаляються, у відповідь на сигнали тканин, що їх оточують, відбувається активація групи генів, що призводять їх до саморуйнування − апоптоз. Апоптоз є одним із проявів принципу самоочищення організму, коли для становлення й збереження цілого багатоклітинний організм жертвує невеликою частиною своїх соматичних клітин. Дійсно, іншим не менш важливим у цьому процесі є захист організму від клітин з безповоротно ушкодженим генетичним апаратом, оскільки в цьому разі виникає небезпека їхнього неконтрольованого росту й загибелі цілого організму. Якщо пошкодження генетичного апарату клітини неможливо відновити, клітина вчиняє самогубство.
Відповідно до центрального постулату молекулярної біології прийнято вважати, що генетична інформація, необхідна для індивідуального розвитку організму, укладена в його генах, які являють собою послідовності нуклеотидів молекул ДНК і РНК. Гени містять інформацію про складові частини організму: сукупності великого числа високо - і низькомолекулярних хімічних сполук, що утворюють його клітини, тканини й органи. При цьому дані про структуру майже всіх низькомолекулярних сполук, що називаються метаболітами, закодовані в генах не прямо, а побічно. Фактично ця інформація є лише програмою біосинтезу метаболітів. Сама ж структура й взаємодія метаболітів один з одним обумовлюються біологічними каталізаторами білкової природи − ферментами, які й здійснюють необхідні взаємоперетворення − метаболізм.
|
|
|
Генетична інформація про структуру білків і ферментів схована в генах не так глибоко. Завдяки існуванню універсального триплетного генетичного коду, послідовності нуклеотидів генів однозначно визначають послідовності амінокислот поліпептидних ланцюгів певних білків. Декодування інформації про структуру білків і нуклеїнових кислот, що супроводжується їхнім біосинтезом, є найважливішим проміжним (але не кінцевим) результатом функціонування (експресії) генів будь-якого організму.
У нормі експресія генів забезпечує існування організму як цілого від початкових до завершальних стадій індивідуального розвитку − від перших поділів стимульованої яйцеклітини до природної смерті організму. Однак, більш широкий підхід до проблеми експресії генів повинен ураховувати не тільки біохімічні наслідки їхньої роботи на молекулярному, надмолекулярному й організменному рівні, але й генетично детерміновані поведінкові реакції груп особин у популяції, а отже, і механізми генетичного контролю розвитку самих популяцій, включаючи цивілізацію. 2.1. Будова та властивості молекули ДНК
Перші дані про хімічні властивості ДНК виникли в 1868 р. На початку 40-х років XXст. було встановлено, що молекула ДНК − це лінійний полімер. Мономерними одиницями її є нуклеотиди, що складаються з азотистих основ, пентози (п'ятивуглеводного цукру) і фосфатної групи (рис. З, а). Фосфатна група приєднана до 5'-атома вуглецю моноцукру, а органічна основа − до 1 '-атома. Основи ДНК бувають двох типів: пуринові [аденін /А/ і гуанін /G/] і піримідинові [цитозин /С/ і тимін /17] (рис. 3, б). У ДНК моноцукор представлено 2'-дезоксирибозою, що містить лише одну гідроксильну групу (ОН), а у РНК − рибозою, яка має дві гідроксильні групи. Нуклеотиди поєднані один з одним фосфодіефірними зв'язками, при цьому фосфатна група 5'-атома вуглецю одного нуклеотиду пов'язана з 3'-ОН-групою дезоксирибози сусіднього нуклеотиду. На одному кінці полінуклеодидного ланцюга є 3'-ОН-група (3'-кінець), а на іншому 5'-фосфатна група (5'-кінець).
У 1953 році Джеймс Уотсон і Френсис Крик на підставі даних рентгеноструктурного аналізу кристалів ДНК зробили висновок, що нативна ДНК складається з двох полінуклеотидних ланцюгів, які створюють між собою спіраль (рис. 4). Полімерні ланцюги з'єднані водневими зв'язками, що утворюються між комплементарними основами А-Т (два зв'язки), G-C (три зв'язки) протилежних ланцюгів (рис. 5). Тому вуглеводно-фосфатний остов молекули має регуляторну структуру. Протилежно цьому послідовність пуринових і піримідинових основ уздовж ланцюга у вищому ступені нерегулярна, кожна молекула ДНК певного типу характеризується власною послідовністю.
Пуринові та піримідинові основи згорнуті усередині подвійної спіралі і розташовані паралельно одна до одної і перпендикулярно до осі спіралі. Два ланцюги утримуються поряд завдяки водневим зв'язкам між парами основ. Аденін завжди спаровується із тиміном, а гуанін − із цитозином. Сувора специфічність спаровування обумовлює комплементарність, тобто взаємну відповідність послідовності основ у двох ланцюгах. Ланцюги молекули ДНК антипаралельні: один з них має напрямок 3'  5', інший − 5' З'. Отже, якщо перевернути спіраль на 180°, то її зовнішній вигляд залишиться незмінним.
5', інший − 5' З'. Отже, якщо перевернути спіраль на 180°, то її зовнішній вигляд залишиться незмінним.

Рис.3. Структура окремого нуклеотиду (а) та нуклеотидів, поєднаних у ланцю (б)

Рис. 5 Основні блоки, що складають дволанцюгову молекулу ДНК
Діаметр спіралі − 20 А (1 ангстрем = 1010 м). Відстань між сусідніми основами вздовж спіралі − 3,4 А, вони повернуті одна відносно другої на 36°, таким чином, на один виток спіралі кожного з ланцюгів припадає 10 нуклеотидів, що відповідає 34 А (рис. 4, а, б).
Довжина дволанцюгової ДНК, як правило, вимірюється числом пар комплементарних нуклеотидів (п.н.). Для молекул ДНК, що складаються з тисяч або мільйонів пар нуклеотидів, прийнято одиниці т.п.н. та м.п.н. відповідно.
Носій генетичної інформації повинен задовольняти дві основні вимоги: відтворюватися (реплікуватися) з високою точністю і детермінувати (кодувати) синтез білкових молекул. Модель ДНК Уотсона-Крика повністю відповідає цим вимогам. По-перше, згідно з принципом комплементарності, кожен ланцюг ДНК може бути матрицею для утворення нового комплементарного ланцюга. Отже, після одного раунду реплікації утворюються дві дочірні молекули, кожна з яких має таку ж нуклеотидну послідовність, як початкова молекула ДНК. По-друге, нуклеотидна послідовність структурного гена однозначно задає амінокислотну послідовність білка, яку вонакодує.
Реплікація. Кожна мономерна одиниця, що приєднується доланцюга,щосинтезується, знаходиться у формідезоксирибонуклеозид-5'-трифосфату (див. рис. 3); фосфатна група, що пов'язана з 5'-вуглеводним атомом дезоксирибози, позначається буквою  , до неї приєднані
, до неї приєднані  -фосфат і далі −
-фосфат і далі −  -фосфат. Під час реплікації - і -фосфатні групи відщеплюються у вигляді пирофосфату, а -фосфатна група поєднується з 3'-ОН-групою останнього нуклеотиду ростучого ланцюга. Синтез ДНК як у прокаріот, так і в еукаріот здійснюється за участю багатьох різних ферментів. Основну роль відіграє ДНК-полімераза, яка послідовно приєднує нові ланки до ростучого полінуклеотидного ланцюга за принципом комплементарності й каталізує утворення фосфодіефірних зв'язків.
-фосфат. Під час реплікації - і -фосфатні групи відщеплюються у вигляді пирофосфату, а -фосфатна група поєднується з 3'-ОН-групою останнього нуклеотиду ростучого ланцюга. Синтез ДНК як у прокаріот, так і в еукаріот здійснюється за участю багатьох різних ферментів. Основну роль відіграє ДНК-полімераза, яка послідовно приєднує нові ланки до ростучого полінуклеотидного ланцюга за принципом комплементарності й каталізує утворення фосфодіефірних зв'язків.
У бактерій реплікація ДНК починається з особливої точки молекули, що називається точкою начала (або сайтом ініціації ) реплікації (оrі, від англ. origin). У ДНК еукаріот існує декілька таких сайтів, і реплікація може починатися в кожному з них. Сегменти еукаріотичної ДНК, що при цьому створюються, скріплюються один з одним за допомогою особливих ферментів. Крім того, в еукаріот є спеціальний фермент теломераза, який поновлює кінці (теломери)хромосом.
Механізм реплікації був запропонований Дж. Уотсоном і Ф. Криком − авторами теорії подвійного ланцюга ДНК. Він передбачає розплетення двох ланцюгів ДНК таким чином, щоб кожен з них міг бути матрицею для зборки іншого ланцюга відповідно до принципу комплементарності. На матричному або батьківському ланцюгу поодинокі нуклеотиди вишиковуються певним чином, а їх наступна полімеризація призводить до утворення нового або дочірнього ланцюгу (репліки), комплементарного першому (рис. 6).
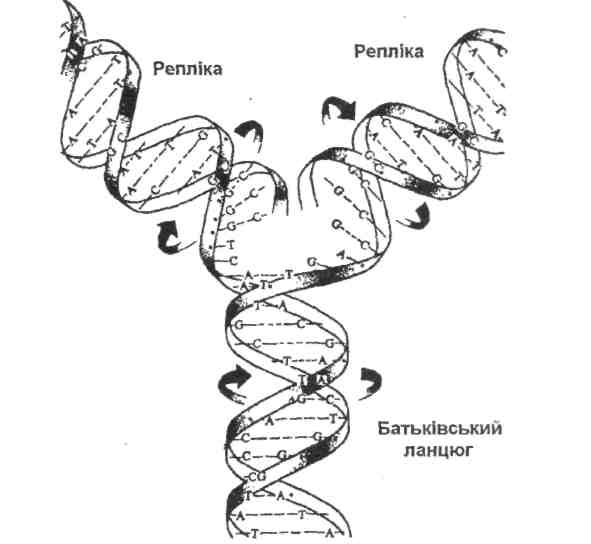
|
Рис. 6. Реплікація молекули ДНК за напівконсервативним механізмом
Запропонований механізм реплікації був названий напівконсервативним, оскільки кожна з ідентичних одна до іншої дочірніх молекул складається з одного старого і одного нового ланцюга ДНК.
Як уже зазначалося, для реплікації необхідне локальне розплетення подвійної спіралі в тій частині молекули, де ДНК в даний момент є матрицею для синтезу дочірніх ланцюгів ДНК. Ця розплетена частина молекули називається реплікативною вилкою.
Реплікація супроводжується пересуванням реплікативної вилки вздовж молекули ДНК і синтезом нових ланцюгів на кожному із старих. Але ланцюги ДНК антипаралельні, і логічно припустити, що один із дочірніх ланцюгів росте у напрямку 5' 3', а інший − в протилежному, 3' 5'.
Оскільки молекула ДНК несиметрична, для реплікації в двох напрямках необхідно два різних ферменти реплікації − дві ДНК-полімерази.
За допомогою однієї ланцюг росте в напрямку 5' 3', і тоді кожен наступний мономер забезпечує сам себе енергією для приєднання до ростучого ланцюга (носієм енергії є трифосфатна група). Такий ріст на полімерному ланцюгу називається «ростом з хвоста». Інша полімераза нарощує ланцюг «з голови», тобто приєднує новий мономер до багатого на енергію 5'-кінця ростучого ланцюга. Цей мономер, у свою чергу, використовує власний трифосфат для приєднання наступного нуклеозидтрифосфату.
Однак, у прокаріот і в еукаріот знайдено, виділено і охарактеризовано лише 5' 3' ДНК-полімерази. Виникає проблема синтезу іншого ланцюга ДНК. Дослідження виявили, що під час реплікації бактеріальної ДНК спочатку на деякий час створюються фрагменти завдовжки 1000...2000 нуклеотидів. Ці фрагменти дістали ім'я їх першовідкривача − фрагменти Оказаки. За реплікації еукаріотичної ДНК довжина фрагментів Оказаки становить 100...200 нуклеотидів. Було також показано, що синтез фрагментів відбувається в напрямку від 5' до 3' кінця, надалі вони з'єднуються у довгі ланцюги ДНК. Так виникло уявлення згідно з яким синтез ДНК здійснюється у напрямку 5' З' (від хвоста до голови). Але на одному з ланцюгів нова ДНК синтезується безперервно, а на іншому −фрагментами, що з'єднуються між собою в єдину полімерну молекулу. Перший з ланцюгів називається лідируючий, а інший -відстаючий. Таким чином, на відстаючому ланцюгу синтез відбувається в напрямку 5' 3', а сам ланцюг росте в напрямку 3' 5'. Основні принципи і механізми реплікації схожі у прокаріот і еукаріот. Найкраще вивчена реплікація у Е. соli. Основним ферментом реплікації у Е. соli є ДНК-полімераза III. Швидкістьсинтезу ДНК за її допомогою становить 1000 нуклеотидів за секунду. Аналогом ДНК-полімерази III в клітинах еукаріот вважають ДНК-полімеразу а (альфа), швидкість реплікації еукаріотичної ДНК у 10 разів повільніша і становить 100 нуклеотидів за секунду.
Механізм реплікації повинен забезпечувати безпомилкове копіювання матриці. Введення в новий ланцюг ДНК некомплементарних нуклеотидів викликає виникнення мутацій, які можуть спричинити генетичні зміни, що порушують структуру і функції генетичного матеріалу. Наслідки таких мутацій відбиваються негативно, а інколи згубно на житті клітини і всього організму.
Для запобігання помилкового парування ДНК-полімерази володіють здатністю до самокорекції. Коригувальний механізм полягає в тому, що перед приєднанням кожного наступного нуклеотиду до ростучого ланцюга, ДНК фермент «перевіряє» правильність парування попереднього нуклеотиду. Якщо нуклеотиди спаровані правильно (за принципом комплементарності), полімераза приєднує наступний нуклеотид, який згодом також буде перевірений. У разі помилки парування, «неправильний» нуклеотид відщеплюється від ланцюга ДНК. Потім перевіряється попередній перед вирізаним нуклеотидом і так далі. Вирізання нуклеотидів здійснюється завдяки тому, що ДНК-полімераза володіє крім полімеразної ще й 3' 5'-екзонуклеазної активністю. Коли полімераза відщепить неспарений нуклеотид або декілька нуклеотидів і дійде до нормально спарених нуклеотидів, відновлюється її полімеразна активність, і синтез ДНК подовжується до виявлення чергової дефектної пари.
Наявність у фермента корегувальної здатності означає, що для ініціації реплікації на матриці йому необхідна хоча б коротка ділянка дволанцюгової ДНК, з якої починається синтез комплементарного ланцюга. Така ділянка називається праймером або запалом.
Для синтезу лідируючого ланцюга, який іде безперервно, праймер необхідний лише на початку полімеразної реакції. Полімераза, що здійснює синтез відстаючого ланцюга, потребує праймера перед синтезом кожного фрагменту. Існує спеціальний фермент, що синтезує праймери. Він називається РНК-праймаза і створює короткі РНК-праймери завдовжки близько 10 нуклеотидів. Після синтезу фрагментів Оказаки праймер треба видаляти.
Для видалення праймерів і забудови створених проломів починає діяти особлива система репарації (відновлення) ДНК. Основна роль у цьому належить ДНК-полімеразі І Е. соli і аналогічним ферментам інших організмів. ДНК-полімеразу 1 можна розглядати як два ферменти на одному поліпептидному ланцюгу. Перший називається фрагментом Кленова і володіє полімеразною і коригувальною (3' 5'-екзонуклеазної) активністю. Інший фрагмент здатний відщеплювати нуклеотиди в напрямку 5' 3', тобто в напрямку синтезу ДНК. Ця 5' 3'-екзонуклеазна активність і надає можливість видаляти праймер.
Синтез ДНК на відстаючому ланцюгу складається, таким чином, з таких етапів:
· РНК-праймаза синтезує праймери на певній (що дорівнює довжині фрагментів Оказаки) відстані однин від одного;
· ДНК-полімера IIIсинтезує фрагменти ДНК, починаючи із 3'-кінця праймера і закінчуючи на 5'-кінці попереднього праймера;
· ДНК-полімераза І подовжує синтез фрагментів ДНК у напрямку 5' 3', одночасно видаляє праймер у тому ж напрямку;
· фрагменти ДНК «скріплюється», тобто створюється фосфодіефірний зв'язок між початком попереднього і кінцем наступного фрагментів.
|
|
|
|
|
Дата добавления: 2017-02-01; Просмотров: 89; Нарушение авторских прав?; Мы поможем в написании вашей работы!