КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Окислительное фосфорилирование
|
|
|
|
Процесс переноса протонов и электронов по дыхательной цепи, локализованной во внутренней мембране митохондрий, к конечному акцептору электронов – молекулярному кислороду – сопровождается очень большим уменьшением свободной энергии. Иначе говоря, передвигаясь от одного переносчика электронов к другому, электроны опускаются на все более низкие энергетические уровни, отдавая порциями свою энергию.
Подсчитано, что при каждом переносе пары электронов с восстановленного пиридинпротеида (с НАД.Н2) на кислород выделяется 52,12 ккал (218,2 кДж) энергии.
При сопоставлении этой величины с величиной стандартной свободной энергии образования АТФ из АДФ и фосфата, равной 7,3 ккал (30,4 кДж) очевидно, что уменьшение свободной энергии при переносе одной пары электронов от НАД.Н2 на кислород достаточно велико для того, чтобы обеспечить возможность образования нескольких молекул АТФ из АДФ и фосфата при условии наличия соответствующего механизма сопряжения фосфорилирования АДФ с окислительным процессом в дыхательной цепи.
При рассмотрении энергетики дыхательной цепи оказалось, что в дыхательной цепи имеются три участка, в которых перенос электронов сопровождается относительно большим изменением стандартной свободной энергии (т.е. высвобождением энергии), превышающим величину стандартной свободной энергии образования АТФ из АДФ и фосфата. Такими участками явились: участок между флавопротеидом и КоQ, участок между цитохромом «в» и цитохромом «с» и участок между цитохромом «а» и цитохромом «а3». Уменьшение свободной энергии на этих участках составляет 9,9-23,8 ккал (40-99,6 кДж), что значительно превышает величину стандартной свободной энергии образования АТФ из АДФ и фосфата, равную 7,3 ккал (30,4 кДж). В других участках дыхательной цепи уменьшение свободной энергии не столь выражено и, по-видимому, не может обеспечить образование молекулы АТФ.
|
|
|
Таким образом, митохондриальная дыхательная цепь напоминает каскадное устройство, поставляющее клетке свободную энергию определёнными порциями.
Идея о наличии сопряжения фосфорилирования АДФ и тканевого дыхания впервые была высказана советским ученым В.А. Энгельгардтом в начале 30-х годов. Впоследствии исследованиями В.А. Белицера, Очоа, Лумис и Липман, Кеннеди и Ленинджера, Митчела, С.Е. Северина, В.П. Скулачева и др. была в значительной мере раскрыта сущность этого процесса.
Установлено, что сопряжёно с передачей протонов и электронов по окислительно-восстановительной цепи ферментов осуществляется важнейший для жизнедеятельности организмов процесс-синтез атф из АДФ и Н3РО4, т.е. высвобождающаяся при тканевом дыхании энергия трансформируется в энергию фосфатной связи АТФ. Этот процесс получил название «окислительное фосфорилирование» и служит для аккумуляции в макроэргических связях АТФ около 40% всей энергии, освобождающейся в процессе тканевого дыхания.
При окислительном фосфорилировании с помощью дыхательной цепи и сопряжено с транспортом протонов и электронов по всей цепи происходит активирование неорганического фосфата и передача его затем на АДФ с образованием АТФ.
Активирование фосфата происходит на описанных выше трех участках митохондриальной дыхательной цепи, характеризующихся повышенным высвобождением свободной энергии.
В случае окисления субстратов пиридиновых дегидрогеназ и восстановленных пиридинпротеидов на каждую пару атомов водорода, поступившую в дыхательную цепь и окислившихся до Н2О, синтезируется три молекулы АТФ, что связано с указанными тремя участками активирования неорганического фосфата и синтезом на каждом из них АТФ из АДФ и активированного фосфата. В случае окисления субстратов флавиновых ферментов (например, янтарной кислоты) и восстановленных флавопротеидов образуется 2 молекулы АТФ, что объясняется выпадением первого участка активирования (участка между флавопротеидом и КоQ).
|
|
|
Величина фосфорилирования выражается показателем эффективности окислительного фосфорилирования, характеризующегося отношением:
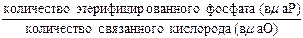
Это отношение, называемое коэффициентом фосфорилирования и обозначаемое как Р/O, оказывается различным в зависимости от подвергающегося окислению субстрата и способа получения митохондрий. Для истинного фосфорилирования, обусловленного реакциями в дыхательной цепи, отношение Р/О равняется 3 (в этом случае происходит окисление восстановленного НАД и субстратов НАД-дегидрогеназы) и 2 (в этом случае происходит окисление восстановленных флавопротеидов и субстратов флавиновых ферментов).
Процесс окислительного фосфорилирования протекает внутри митохондрий – субклеточных частичках специфического строения. В клетках их может находиться от нескольких сотен до нескольких десятков тысяч. Характерной особенностью строения митохондрий является наличие у них двух мембран, из которых внутренняя имеет большую протяженность и образует выпячивания (кристы), погруженные во внутреннее основное вещество митохондрий, называемое матриксом. Толщина наружной мембраны составляет приблизительно 7,0 нм, а внутренней мембраны – 5,0-5,5 нм.
Внутренняя поверхность внутренней мембраны покрыта расположенными в определенном порядке частицами сферической формы (диаметром 8,0-9,0 нм), получивших название элементарных структурных единиц.
Мембраны состоят из липидов (1/3 часть) и белков (2/3 части), матрикс представляет собой студнеобразную, полужидкую массу, состоящую приблизительно на 50% из белка. Около 20-25% общего белка внутренней мембраны составляют белки ферментов, участвующих в образовании дыхательной цепи и окислительного фосфорилирования, тогда как остальные белки являются структурными.
Дыхательные ансамбли, состоящие из флавопротеидов, убихинона, железосерных белков и цитохромов, располагаются в плоскости внутренней мембраны митохондрий в основании элементарных структурных единиц, которые представляют собой, по современным воззрениям, АТФ-азную систему (АТФ‑синтетазу), включающую два особых белковых комплекса F1 – F0 (факторы сопряжения), обеспечивающие фосфорилирование АДФ в АТФ в процессе переноса электронов по дыхательной цепи. Дыхательные ансамбли равномерно распределены по плоскости внутренней мембраны. F0-комплекс (фактор) состоит из 4-х субъединиц, которые пронизывают мембрану поперек, ответственен за перенос Н+. F1-комплекс (фактор) связан с F0 и состоит из 9-10 субъединиц 5-ти типов, образует грибовидный выступ на мембране, непосредственно ответственнен за синтез АТФ.
|
|
|
Внутри митохондрий, кроме ферментов дыхательной цепи и окислительного фосфорилирования, находятся также ферменты лимоннокислого цикла, окислительного декарбоксилирования пировиноградной кислоты, β‑окисления жирных кислот, орнитинового цикла и др.
Реакции лимоннокислого цикла, процессы переноса электронов по дыхательной цепи и окислительного фосфорилирования происходят внутри митохондрий или на внутренней поверхности внутренней мембраны. Поэтому молекулы фосфата, АДФ, субстраты лимоннокислого цикла и тканевого дыхания, прежде чем подвергнуться окислению, должны сначала проникнуть внутрь митохондрий. Однако внутренняя мембрана непроницаема для катионов Na+, К+, Мg+, анионов С1-, Вr-, NO3-, Н+, сахаров (таких как сахароза), большинства аминокислот, окисленных и восстановленных НАД и НАДФ, нуклеозид-5-моно-, ди- и трифосфатов (в том числе АДФ и АТФ), коэнзима А и его эфиров.
Внутренняя мембрана проницаема только для воды, для небольших нейтральных молекул, таких как мочевина и глицерин, для жирных кислот с короткой цепью.
Оказалось, что для переноса специфических метаболитов через мембрану внутренняя мембрана содержит несколько ферментоподобных соединений (пермеаз или транслоказ). Такие переносчики идентифицированы для АДФ и АТФ, для фосфата и для некоторых промежуточных продуктов лимоннокислого цикла (сукцината, малата, изоцитрата, цитрата, цис-аконитата), а также для глутамата и аспартата.
|
|
|
Благодаря этим переносчикам осуществляется сложный двусторонний обмен промежуточными продуктами лимоннокислого цикла, фосфатом, АДФ и АТФ между цитоплазмой и внутренним отделением митохондрий. В частности, благодаря функционированию АДФ‑АТФ‑переносчика, требуемое для окислительного фосфорилирования количество АДФ входит внутрь митохондрий через внутреннюю мембрану, а одновременно эквимолекулярное количество АТФ выходит из нее в цитоплазму.
Протоны и электроны цитоплазматического НАД.Н2 (образовавшегося, к примеру, на окислительной стадии расщепления глюкозы в цитоплазме) могут поступать внутрь митохондрий непрямым путем, без переноса самих молекул НАД. Это осуществляется с помощью глицерофосфатного или малатного челночного механизма. Предполагают, что существует также лактатный челночный механизм.
Аналогичный механизм осуществляется для перемещения протонов и электронов из митохондрий в цитоплазму (что происходит, к примеру, при биосинтезе глюкозы из пирувата в цитоплазме).
Сущность глицерофосфатного челночного механизма сводится к следующему. Цитоплазматический НАД.Н2 сначала реагирует с цитоплазматическим фосфодиоксиацетоном (одним из промежуточных продуктов гликолиза), образуя глицерофосфат. Эта реакция катализируется НАД-зависимой цитоплазматической глицерофосфатдегидрогеназой.
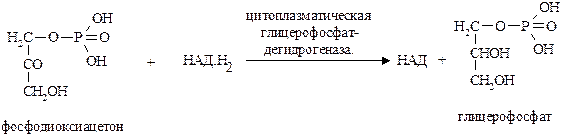 |
Образовавшийся глицерофосфат способен легко проникать через митохондриальные мембраны внутрь митохондрий, где внутримитохондриальная флавин-зависимая глицерофосфатдегидрогеназа снова окисляет глицерофосфат до фосфодиоксиацетона:
Восстановленный флавопротеид вводит приобретенные им электроны в дыхательную цепь(наКоQ), обеспечивая окислительное фосфорилирование двух молекул АДФ, а фосфодиоксиацетон выходит из митохондрий в цитоплазму, где снова может служит акцептором электронов для новой молекулы цитоплазматического НАД.Н2. Высказывается также мнение: окисление ФП.Н2 не приводит к образованию 2 молекул АТФ, а приводит к высвобождению энергии в виде тепла.

Малатный челночный механизм включает систему промежуточных соединений: оксалоацетат – малат. Цитоплазматический НАД.Н2 сначала реагирует с оксалоацетатом при участии цитоплазматической малатдегидрогеназы, образовавшийся малат с помощью транслоказы переносится внутрь митохондрий, где под влиянием митохондриальной дегидрогеназы дегидрируется. Образовавшийся НАД.Н2 окисляется флавопротеидом дыхательной цепи митохондрий, путем окислительного фосфорилирования образуется три молекулы АТФ. Полагают, что малатный челночный механизм является наиболее активным механизмом переноса восстановительных эквивалентов из цитоплазмы в митохондрии.
Для объяснения механизма окислительного фосфорилирования существует три гипотезы, а именно: гипотеза химического сопряжения, гипотеза хеми-осмотического сопряжения и гипотеза механохимического или конформационного сопряжения окисления и фосфорилирования.
В настоящее время наиболее серьезное обоснование получила гипотеза хеми-осмотического сопряжения, предложенная Митчелом в 1961 году и получившая развитие в исследованиях советского ученого В.П. Скулачева (1972 г.). В 1978 г. Митчелу за разработку хеми-осмотической гипотезы была присуждена Нобелевская премия.
Исходя из того, что митохондриальная мембрана является существенным элементом механизма окислительного фосфорилирования и что она непроницаема для ионов водорода (Н+), согласно хемиосмотической гипотезы предполагается, что при тканевом дыхании в процессе движения электронов вдоль дыхательной цепи каждая пара электронов, поставляемая НАД.Н2, трижды пересекает внутреннюю мембрану митохондрий, каждый раз захватывая пару протонов из матрикса и в итоге переносит три пары протонов из внутренней части митохондрий (матрикс) через мембрану наружу (в межмембранное пространство). В результате транслокации протонов через мембрану на внутренней мембране возникает протонный градиент (электронно-химический потенциал), представляющий собой форму запасания свободной энергии. Общая энергия протонного градиента складывается из концентрационного (или осмотического) компонента, определяемого разницей рН по обеим сторонам мембраны, и электрического компонента, обусловленного движением положительно заряженных протонов через мембрану (разница рН примерно =1,4 единицы, электрический потенциал – около 140 мВ.). Из-за разницы в концентрационном и электрическом потенциале протоны, вынесенные из митохондрий, стремятся пересечь мембрану в обратном направлении. Обратное движение протонов через мембрану (через протонные каналы – белок Fо) под влиянием протонного градиента сопряжено с фосфорилированием, или иначе способно привести к работе, такой как фосфорилирование: АДФ+Ф®АТФ+НОН. Предполагается, что два протона, переносимые с помощью белкового комплекса Fо через мембрану (через протонный канал), взаимодействуют с одним из кислородов (или ОН-групп) фосфата, связанного с белковым комплексом F1 ферментной системы F1 – F0 АТФ‑синтетазы, что приводит к высвобождению кислорода с образованием воды и делает фосфатную группу высокореактивной и способной связываться с АДФ с образованием АТФ. Установлено, что на каждые два протона, пересекающих комплекс F1 – F0 образуется одна молекула АТФ из АДФ и активированного описанным способом неорганического фосфата.
Характерная особенность рассмотренной хеми-осмотической гипотезы состоит в том, что образование АТФ в процессе окислительного фосфорилирования происходит без участия высокоэнергетических промежуточных продуктов. Роль промежуточного звена, движущей силой процесса служит электрохимический потенциал (протонный градиент), возникающий на мембране митохондрий за счет энергии, выделяемой в процессе переноса электронов по дыхательной цепи. Согласно наблюдениям В.П. Скулачева, в процессе дыхания на мембране митохондрий, хлоропластов и бактерий действительно возникает мембранный потенциал, достаточный для энергетического обеспечения реакции синтеза АТФ из АДФ и фосфата.
Не следует думать, что любое окисление органических соединений в живых организмах сопряжено с фосфорилированием, так же как и фосфорилирование не обязательно должно быть окислительным. В настоящее время известно несколько сот реакций окисления, но менее десятка их сопряжено с одновременным активированием неорганического фосфата.
Кроме окислительного фосфорилирования различают также субстратное, фотосинтетическое (растения), хемосинтетическое (микроорганизмы) фосфорилирование.
Объем окислительного фосфорилирования в значительной мере зависит от проницаемости мембраны митохондрий. Митохондриальная мембрана может пропускать внутрь митохондрий посредством глицерофосфатного или малатного челночного механизма большее или меньшее количество восстановительных эквивалентов с цитоплазматической НАД.Н2, которые включаются здесь в процессы окислительного фосфорилирования, тогда как на поверхности митохондрий в цитоплазме НАД.Н2 окисляется свободно, при этом не образуется АТФ, а выделяющаяся энергия превращается в тепло. Однако и внутри митохондрий может произойти переключение с окисления, сопряженного с фосфорилированием, на свободное окисление (и обратно), сопровождаемое образованием тепла. Это происходит при разобщении переноса электронов и фосфорилирования в дыхательной цепи, что может произойти, в частности, при понижении температуры или воздействии некоторых химических липофильных веществ (замещенных фенолов, к примеру, 2,4-динитрофенола, а так же свободных жирных кислот, фенилгидразонов, грамицидина, арсената, дикумарола, тироксина и др.). Их назвали ионофорами. Следует заметить, что действие химических разобщителей (так называемых протонофоров или ионофоров), способных нивелировать протонный градиент путем переноса протонов через мембрану митохондрий в обратном направлении – в матрикс, минуя протонный канал, послужило важным доказательством хеми-осмотической гипотезы сопряжения фосфорилирования и тканевого дыхания. Значительные исследования в этой области были выполнены В.П. Скулачёвым.
Интенсивность окислительного фосфорилирования регулируется соотношением в клетке содержания АТФ, с одной стороны, и АДФ и неорганического фосфата, с другой. Причем, два последних вещества активируют процесс окислительного фосфорилирования. При усилении распада АТФ на АДФ и Н3РО4 в процессе реакций, идущих с потреблением энергии, и накопления последних в клеточном содержимом, автоматически усиливается окислительное фосфорилирование, т.е. биосинтез АТФ.
|
|
|
|
Дата добавления: 2014-01-05; Просмотров: 4574; Нарушение авторских прав?; Мы поможем в написании вашей работы!