КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Кормление и питание рыб
|
|
|
|
6.1.Теоретические основы питания рыб
6.1.1.Парадигмы питания
Наблюдения, касающиеся закономерностей питания животных, относятся к глубокой древности.
Первую научную парадигму питания принято связывать с достижениями античных натурфилософов. Согласно представлениям Аристотеля, Галена и их последователей, пища, поступающая в желудочно-кишечный тракт животных, в результате сложных процессов неизвестной природы преобразуется в кровь, которая и обеспечивает питание всего организма.
Вторая классическая парадигма питания начала формироваться в конце XVIII в. одновременно с развитием аналитических методов исследования. Главным достижением основополагающих экспериментов Л. Спалланцани, Р. Реомюра и А. Лавуазье было понимание необходимости растворения пищи перед поступлением ее во внутреннюю среду организма. В результате выдающихся достижений химии, физиологии и энзимологии уже во второй половине XIX в. были сформулированы основные принципы, позволившие разработать теорию сбалансированного питания. Согласно этой теории, в организм животных должны поступать пищевые вещества,в количественном и качественном отношении близкие веществам, израсходованным в процессе жизнедеятельности. При этом начальные этапы ассимиляции пищи осуществляются за счет двух типов пищеварения (полостного и внутриклеточного) и последующего всасывания полезных пищевых веществ - нутриентов. Баластные вещества относились к нежелательным, как и вредные вещества пищи.
В конце XIX—начале XX в. появились представления о необходимости усовершенствования не только технологии питания, но и жизнедеятельности организма. В частности, возникла идея отбрасывания балластных веществ и формирования максимально обогащенной оптимизированной пищи, состоящей из нутриентов. Многие ученые до последнего времени полагали, что можно создать идеальную пищу, которая в виде питательных смесей вводилась бы в желудочно-кишечный тракт.
|
|
|
В рамках классической парадигмы питания к середине XX в. были описаны важнейшие закономерности, касающиеся всех звеньев сложного процесса экзотрофии. Вместе с тем в последующие десятилетия в различных областях биологии, в том числе и в науке о питании, были сделаны столь важные открытия, что потребовался пересмотр многих представлений.
На базе классических концепций и достижений современной науки в настоящее время формируется новая, третья по счету, парадигма питания.
Одним из основных достижений классической парадигмы питания являлось осознание главной роли не только энергетически емких основных компонентов пищи, таких как белки, жиры и углеводы, но и таких как витамины, гормоны, нуклеиновые кислоты и др. Особое внимание при этом уделялось биохимическому элементному составу пищи, в частности соотношению заменимых и незаменимых аминокислот, насыщенных и ненасыщенных жирных кислот, различных гексоз и пентоз, витаминов, минеральных солей и т. д. Согласно данной парадигме, нутриенты, поступающие в пищеварительный тракт в форме полимеров, в процессе пищеварения гидролизуются до уровня мономеров главным образом за счет механизма полостного пищеварения. При этом схема потоков различных веществ из желудочно-кишечного тракта во внутреннюю среду организма представлялась исключительно простой (рис. 1).
Предполагалось, что роль балластных веществ, токсинов и микрофлоры в будущем должна снижаться за счет создания элементных диет, содержащих нутриенты в форме мономеров. На базе этих представлений разрабатывались и продолжают разрабатываться многочисленные диеты для человека и рационы для домашних животных.
|
|
|
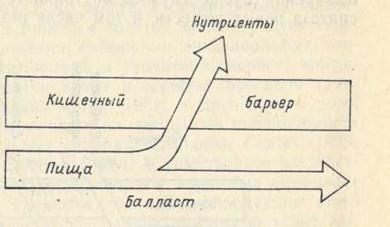
Рис. 1. Схема потоков веществ из желудочно-кишечного тракта во внутреннюю среду организма в соответствии с классической парадигмой
(по: Уголев, 1984).
Однако, несмотря на важную роль теории сбалансированного питания, в научном обосновании пищевых потребностей человека и животных в настоящее время не вызывает сомнения ошибочность некоторых старых представлений.
Более того, балансный подход и вытекающая из него идея рафинированной (безбалластной) пищи, по-видимому, принесли определенный вред. Действительно, широкое использование рафинированной пищи способствовало развитию так называемых болезней цивилизации, таких как ожирение, гипертония, атеросклероз, инфаркт миокарда, диабет, гастрит, холецистит, рак, парадонтоз и т. д. (Haenel, 1979, 1980; Уголев, 1985, и др.)
Эти факты, наряду с открытием новых механизмов пищеварения и транспорта, углублением представлений об энтериновой гормональной системе, анализом различных характеристик безмикробных и контрольных животных, а также данными по влиянию элементных диет на различные системы организма человека и животных обусловили пересмотр ряда основополагающих положений теории сбалансированного питания и классической парадигмы питания в целом.
В связи с современными достижениями науки в этой области в настоящее время формируется новая парадигма питания, центральное положение в которой занимает теория адекватного питания, включающая в себя как важную составную часть теорию сбалансированного питания. Эта теория содержит ряд новых основополагающих постулатов:
1) питание поддерживает молекулярный состав организма и возмещает его энергетические и пластические расходы на основной обмен, внешнюю работу и рост (это положение является общим для классической и новой теорий питания);
2) необходимыми компонентами пищи служат не только нутри-енты, но и балластные вещества;
3) нормальное питание обусловлено не одним потоком нутриентов из желудочно-кишечного тракта, а несколькими потоками нутритивных и регуляторных веществ, имеющих жизненно важное значение;
4) в метаболическом и особенно трофическом отношениях ассимилирующий организм рассматривается как надорганизм;
|
|
|
5) существует эндоэкология организма - хозяина, образуемая микрофлорой и простейшими кишечника;
6) баланс пищевых веществ достигается в результате освобождения нутриентов из структур пищи при ферментативном расщеплении ее макромолекул за счет полостного и мембранного пищеварения (в ряде случаев внутриклеточного), а также вследствие синтеза новых веществ, в том числе незаменимых (Уголев, 1987).
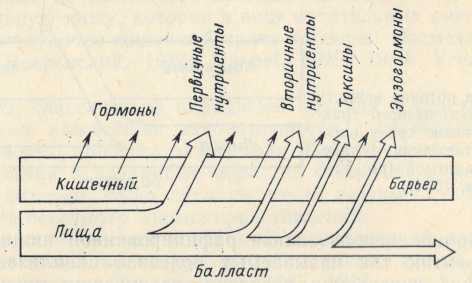
Рис. 2. Схема потоков веществиз желудочно-кишечного тракта во внутреннюю среду организма в соответствии с новой парадигмой (по: Уголев, 1984).
Рассмотрим более подробно некоторые новые положения этой теории.
· Согласно современным представлениям, помимо основного потока нутриентов, учитываемого теорией сбалансированного питания, во внутреннюю среду организма из пищеварительного тракта поступает еще ряд важных потоков (рис. 2). Прежде всего, это поток вторичных нутриентов, образующихся из балластных веществ в результате деятельности микрофлоры, в состав которого входят витамины, незаменимые аминокислоты, углеводы, жиры и т. д. Кроме того, существуют потоки нутриентов, модифицированных микрофлорой, а также поток продуктов жизнедеятельности бактерий (Caspary et al., 1981). Исключительно важную роль играют потоки гормонов и других физиологически активных веществ. Известно, что эндокринные клетки желудочно-кишечного тракта продуцируют около 30 гормонов и гормоноподобных субстанций, контролирующих не только функции пищеварительного аппарата, но и важнейшие эндокринные и метаболические функции всего организма (Уголев, 1978; Larsson, 1979; Климов, 1983, и др.). Предполагается, что эндокринная система пищеварительного аппарата наряду с этим участвует в регуляции аппетита (Уголев, 1978). В последние годы появились сведения о продуцировании клетками тонкой кишки эндорфинов и энкефалинов, а также об образовании при гидролизе молока и пшеницы морфиноподобных веществ, получивших название -экзорфинов (Chang et al., 1981; Hazum et al, 1981). Эти данные свидетельствуют о существовании потоков как эндогенных, таки экзогенных гормонов и других физиологически активных веществ.
|
|
|
Особую роль в формировании структурных и ряда функциональных характеристик пищеварительного тракта играет поток бактериальных метаболитов. В частности, у безмикробных животных снижена масса стенки тонкой и толстой кишок, редуцирован эпителий, снижена митотическая активность эпителиальных клеток, недоразвита иммунная система. Бактериальные метаболиты влияют на кишечную проницаемость и активность пищеварительных гидролаз. Существенное воздействие на пищеварительную и другие системы животных оказывает гистамин бактериального происхождения. Гиперпродукция этого амина, образующегося при декарбоксилировании аминокислот, вызывает язвы желудка, аллергии, повышенную чувствительность к нарушению гипоталамо-гипофизарных функций.
В связи с усилением антропогенного загрязнения среды обитания увеличился поток веществ, попадающих в желудочно-кишечный тракт человекаи животных с водой и пищей (Уголев, 1985). Таким образом, во внутреннюю среду организма поступает не один, а семь или более энтеральных потоков различных веществ, которые участвуют как в обмене веществ, так и в регуляции метаболизма. В свете этих представлений балластные вещества являются не только полезными, но и необходимыми компонентами пищи, так как служат основой для продукции кишечной микрофлорой исключительно важных веществ, способных компенсировать недостающие компоненты. Таким образом, в метаболическом отношении все животные представляют собой надорганизмы. Поскольку наличие кишечной микрофлоры является эволюционно закрепленной формой существования большинства животных, сложные взаимоотношения между микрофлорой и макроорганизмом необходимо учитывать при оценке различных аспектов жизнедеятельности животных.
Новая парадигма питания и ассимиляции пищи придает большое значение защитным функциям пищеварительного тракта. Как известно, с пищей в организм поступают аллергены и токсины. Попаданию многих из них во внутреннюю среду препятствует, прежде всего, ограниченная проницаемость эпителия для макромолекул, в число которых входят белки, мукополисахариды и другие соединения, обладающие антигенными свойствами. Кроме того, в тонкой кишке функционирует иммунная система, представленная пейеровыми бляшками, содержащими В-, М- и Т-клетки.
Помимо этого существует мощный лейкоцитарный слой, причем некоторая часть лимфоцитов обнаружена внутри энтероцитов. Наконец, на поверхности энтероцитов обнаружены иммуноглобулины А и Е, создающие в области гликокаликса дополнительный защитный слой. Описаны также защитные функции внутриклеточных пептидази системы антитоксических реакций, и значительной мере связанные с активностью цитохромов Р450.
Эти данные свидетельствуют о существовании разнообразных защитных механизмов в пищеварительномтракте, который наряду с печеночным барьером выполняет важные функции по устранению антигенной и токсической агрессий.
Вместес тем в настоящее время признается, что небольшие дозы токсинов и аллергеновнеобходимы для поддержания пищеварительной и иммунной системы организма в функционально активном состоянии.
Выше были перечислены основные положения теории адекватного питания, касающиеся организменного уровня организации материи. Однако новая парадигма питания охватывает все звенья трофических отношений, включая популяционный, биоценотический и биосферный.
С этих позиций необходимо охарактеризовать основные классификации типов питания животных.
6.1.2. Роль микрофлоры в гидролизе и трансформации пищевых субстратов
В последние годы усилился интерес к микрофлоре пищеварительного тракта рыб. Благодаря этим исследованиям утвердились представления о том, что микрофлора кишечника необходима для нормального роста и развития рыб. Причем взаимоотношения макро- и микроорганизмов кишечника рассматриваются с позиций облигатного симбиоза. В монографии В. Н. Лубянскене с соавторами (Лубянскене и др., 1989) анализируется поступление микроорганизмов в кишечник рыб, роль микрофлоры, как поставщика ферментов, а также в трансформации органических веществ первичной пищи в белки и другие компоненты их собственного организма.
К наиболее важным результатам этой работы относятся:
· Общее количество бактерий в пищеварительном тракте находится в прямой зависимости от интенсивности питания рыб. Наибольшее их количество установлено в июле—августе, когда наблюдается и самое интенсивное питание рыб.
· Значительное влияние на микрофлору оказывает состав пищи. Так, наибольшее число бактерий в 1 г содержимого кишечника (1,29- 1012 кл.) отмечено в период интенсивного питания в пищеварительном тракте линей, выросших на естественной пище, значительно меньше их в кишечниках белого амура и линя, выращенных на комбикорме.
· На примере карпа установлено, что с возрастом число бактерий в пищеварительном тракте рыб увеличивается.
· При исследовании различных видов рыб (карп, белый амур, линь, карась) установлено, что выделенные из пищеварительного тракта бактерии принадлежат к родам Pseudomonas, Micrococcus, Bacillus, Bacterium, Pseudobacterium, Azotobacter, Sarcina, среди которых явно доминируют представители рода Pseudomonas. При этом известно, что различные виды бактерий, принадлежащих к роду Pseudomonas, характеризуются высокой протеолитической активностью, в том числе активностью внеклеточных и внутриклеточных гидролаз. Есть сведения о нейтральной и щелочной протеиназах, а также эластазе.
· При изучении секреции свободных аминокислот микрофлорой (в основном рода Pseudomanas) в кишечнике рыб (карп, белый амур и линь) установлено, что она способна синтезировать до 13 свободных аминокислот: лизин, аргинин, гистидин, аспарагиновую кислоту, треонин, аланин, метионин, валин, фенилаланин, лейцин, серии и глютаминовую кислоту.
· Продукция свободных аминокислот кишечными микроорганизмами изменяется в зависимости от возраста рыб: наибольшая у сеголеток, меньшая — у двухлеток, минимальная— у трехлеток. Количество общих свободных аминокислот и отдельных аминокислот в обменном фонде клетки меняется в течение ее развития.
· При различных условиях питания и при голодании рыб кишечные микроорганизмы секретируют до 17 свободных аминокислот (кроме указанных выше цистеин, аспарагин, тирозин и триптофан). Кроме того, установлена важная роль микрофлоры в фиксации молекулярного азота. В условиях интенсивного питания рыб самая высокая нитрогеназная активность во всем пищеварительном тракте установлена у толстолобика — до 3,648 нмоль С2Н4/ (мг белка • ч).
· У карпа, леща и плотвы при интенсивном питании в природных условиях в пищеварительном тракте азотфиксация происходит слабее, чем у растительноядного толстолобика, но сильнее чем у хищника щуки.
· Велика роль бактерий и в продуцировании витаминов. Так, до 50% потребляемых рыбами витаминов продуцируется микрофлорой их пищеварительного тракта. Кроме того, существенна роль бактерий в синтезе антибиотических веществ, подавляющих рост патогенной микрофлоры.
Вышеприведенные данные свидетельствуют о том, что у рыб, как и у других животных, микрофлора пищеварительного тракта играет исключительно важную роль в деполимеризации и трансформации первичной пищи. Это обстоятельство дает основание считать, что у рыб существует вторичный поток нутриентов. Последнее имеет большое значение для анализа механизмов пищеварения и эффективности питания рыб из естественных ихтиоценозов, а также для контроля и коррекции питания прудовых популяций рыб.
6.1.3. Типы питания животных
Ранее вся биота планеты по типу питания делилась на 2 группы:
- автотрофы, использующие в качестве пищевых субстратовнеорганические вещества,
- гетеротрофы, потребляющие кроме неорганических органические вещества, синтезируемые преимущественно автотрофами.
Эта широко распространенная классификация в настоящее время дополняется. В последние годы были предложены классификации, учитывающие источник энергии или физическое состояние пищи (Prosser, 1977), источник энергии, а также источник углерода, азота и других органических и неорганических соединений (Stanier et al., 1979a—1979c).
Согласно Станиеру и соавторам (Stanier et al., 1979a—1979с), существуют 4 группы организмов:
- фотоавтотрофы (водоросли, высшие растения и многие фотосинтезирующие бактерии), использующие свет как источник энергии, а СО2 как источник углерода;
- фотогетеротрофы (пурпурные и зеленые бактерии), использующие свет в качестве энергии и какое-либо органическое вещество в качестве источника углерода;
- хемоавтотрофы, использующие химический источник энергии, освобождаемой при окислении ряда восстановленных неорганических соединений (NH3, NO2, H2), восстановленных форм серы (H2S, S, S2O3), или закисное железо (только бактерии). Эти организмы иногда называют хемолитотрофами, так как они способны к росту в минеральной среде в отсутствие источника света;
- хемогетеротрофы (все многоклеточные животные, простейшие, грибы и подавляющее большинство бактерий), использующие химический источник энергии и органические вещества в качестве источника углерода.
Поскольку корректное определение процессов питания подразумевает ассимиляцию всех веществ, поступающих извне, а живые организмы являются экзотрофами и, следовательно гетеротрофами, была предложена натуральная классификация биоты (Уголев, 1980). Согласно ей все живые организмы распределяются по шкале, на одном полюсе которой находятся полные абиотрофы, на другом — полные биотрофы. В настоящее время полных абиотрофов практически не существует (известны лишь литотрофы). Полная биотрофия присуща некоторым монофагам, симбионтам и эмбрионам. Хищники близки к полным биотрофам, однако в их пищу входят соли неорганического происхождения и вода. К биотрофии относится также эндотрофия в период голодания животных и темнового питания растений.
Натуральная классификация оказывается исключительно полезной при анализе трофических цепей, в частности перехода от одного участка трофической цепи к другому.
Важно отметить, что биотрофия существует в нескольких вариантах:
- естественная, или витальная,
- экзотрофия (питание живыми организмами и их элементами),
- поствитальная биотрофия,
- сапрофитизм (питание продуктами жизнедеятельности других организмов),
- эндотрофия.
Способы питания различных животных могут существенно различаться. В связи с этим сохраняет свое значение традиционное деление животных по типу питания, базирующееся на анализе спектра кормовых объектов, в частности моно- и эврифагов, фито- и зоофагов, хищников и «мирных», бенто- и планктофагов и т. д.
Вместе с тем в процессе эволюции сложилось не только принципиальное сходство животных разных видов по молекулярному составу, но и сходство систем, осуществляющих подготовку сложных органических, как правило, высокомолекулярных соединений к последующей ассимиляции. Эти системы состоят из многочисленных гидролаз, разрушающих биополимеры до уровня моно- и олигомеров, пригодных к всасыванию и ассимиляции.
6.1.4. Трофические цепи и биогеоценозы
В настоящее время принято считать, что синтезы различных биологических веществ возникали в процессе эволюции постепенно, по мере выживания организмов, способных осуществлять те или иные процессы. Построение пищевой цепи, по-видимому, базировалось на редуцентах и завершалось фото- и минерал-зависимыми абиотрофами, так как системы фото- и хемосинтеза, необходимые для абиотрофии, в эволюционном отношении являются сравнительно молодыми.
Предполагается, что первичные бионты получали основные органические материалы в виде мономеров из небиологических источников. При этом первичная гетеротрофия возникла на основе использования молекул, обеспечивающих функции внутриклеточного гидролиза, связанного с перестройкой и мобилизацией собственных полимерных структур. Эти древние гидролазы, служившие основой для формирования эндотрофии, впоследствии стали использоваться для реализации процессов экзотрофии.
Древность происхождения и первичность эндотрофии позволяют понять сходство механизмов эндо- и экзотрофии у столь значительно различающихся по уровню филогенетического развития организмов, как бактерии, высшие растения, беспозвоночные и позвоночные животные.
Благодаря принципиальному сходству молекулярного состава и механизмов усвоения пищевых веществ с помощью ферментативных систем, обеспечивающих деполимеризацию органических соединений, оказывается возможным изменение не только спектра питания, но и места животных в трофической цепи. Это фундаментальное свойство живых систем позволяет им адаптироваться к изменению условий питания и тем самым полнее использовать кормовую базу. Это же свойство позволяет видам, относящимся к различным типам и даже царствам, занимать аналогичные трофические ниши.
Одним из наиболее важных для биологии выводов новой парадигмы питания является то, что процветание любого вида в значительной мере определяется его положением в трофической цепи. Иными словами, существование отдельных видов и популяций зависит не только от наличия пищи, но и от их способности быть источником питания для следующих звеньев трофической цепи. Этот, на первый взгляд парадоксальный, вывод был подготовлен многочисленными данными экологических работ, свидетельствующих о том, что главной побудительной причиной активности всех животных является необходимость поиска пищи, количество, качество и доступность которой влияют как на распределение, так и на численность различных популяций животных.
Полезными для анализа трофических отношений различных животных оказались понятия фагичности и трофичности организмов.
Фагичностьорганизмов — доступность объектов питания для потребляющих их консументов,
Трофичность организмов — питательность и способность быть ассимилированными.
Именно сочетание фагичности и трофичности видов обеспечивает их процветание, а также взаимные адаптации трофических партнеров. Из экологической литературы хорошо известно, что численность и другие характеристики популяций хищника и жертвы находятся в строгой взаимозависимости. Так, уменьшение численности популяции жертвы может вызывать уменьшение или даже исчезновение популяции хищника, в той или иной мере отражающееся на состоянии популяций консументов более высокого порядка.
При благоприятных условиях питания численность популяции потенциальной жертвы может увеличиваться и вызвать увеличение популяции хищника. Однако при значительном увеличении популяции хищника возможны нарушение структуры и снижение темпа воспроизводства популяции жертвы, которые приводят к депрессии популяции хищника. Эти факты свидетельствуют о важной роли не только источника пищи, но и механизма обратной связи, регулирующего состав и численность животных, входящих в экосистему. Не вызывает сомнения высокая эффективность такой регуляции, а также вытекающая из этого необходимость крайне осторожного воздействия на различные экосистемы извне.
Существуют многочисленные примеры печальных последствий вмешательства человека в функционирование экосистем, когда поголовье хищников истреблялось или искусственно увеличивалось в результате специальных мероприятий. Кроме того, описаны механизмы регуляции трофических отношений животных, заключающиеся в изменении интенсивности питания хищников. Так, офиуры и морские звезды, питающиеся пластиножаберными моллюсками, голодают в течение 1—2 мес, пока личинки объектов питания не увеличат массу на 2—3 порядка, и таким образом сохраняют популяцию жертвы. Исключительный интерес в этом плане представляют взаимоотношения паразит—хозяин, которые в процессе эволюции приобрели более нейтральные формы и приблизились в ряде случаев к симбионтным отношениям. Ясно, что уменьшение вредных для хозяина последствий паразитизма приводит к улучшению состояния его популяции и тем самым способствует сохранению паразитов.
Следует подчеркнуть схематичность описанных выше примеров трофических отношений животных. В естественных условиях, когда в состав экосистем входят многочисленные эврифаги, трофические взаимоотношения животных представляют собой не трофические цепи, а трофические сети. При этом и фагичность, и трофичность объектов питания зависят не только от биотических, но и абиотических факторов среды. Так, доступность эктотермных животных для гомойотермных хищников увеличивается по мере снижения температуры окружающей среды. Интенсивность процессов экзотрофии у пойкилотермных хищников при уменьшении температуры, напротив, снижается. Значительное влияние на фагичность и трофичность жертвы оказывает также газовый состав среды обитания, давление, доступность и качество воды, состав почв, грунтов и т. д. Это делает понятной значительную зависимость трофических отношений различных животных от структуры биогеоценозов.
Таким образом, в основе трофических взаимоотношений животных лежит принципиальное сходство молекулярного состава биоты, а также единство механизмов экзотрофии. Трофические цепи, или сети, большинства современных организмов исключительно сложны и в значительной мере зависят не только от биотических, но и абиотических компонентов биогеоценозов.
Несмотря на то, что носителями жизни являются отдельные организмы, а универсальным элементом — клетка, жизнь на Земле возможна лишь как планетарное явление. Необходимым условием сохранения жизни в планетарном масштабе является поддержание равновесия между скоростью синтеза и деструкции вещества. Известно, что ежегодно на Земле образуется приблизительно 230 млрд тонн органических веществ, которые разрушаются различными биотрофами. При этом в каждом следующем трофическом звене масса органического вещества уменьшается, как правило, на порядок.
Для понимания трофических взаимоотношений организмов в планетарном масштабе исключительно полезной оказалась концепция биосферы. Как известно, значение этого термина существенно изменилось под влиянием работ Зюсса и особенно представлений В. И. Вернадского. Согласно Зюссу, биосфера — особая оболочка Земли, образованная живыми организмами. Большой заслугой Вернадского было не только введение в понятие биосферы области распространения жизни на Земле (населенную организмами поверхность суши, толщу вод и нижнюю часть атмосферы), а также включение наряду с организмами среды их обитания, но и постулирование изменения в процессе эволюции как биотических, так и абиотических компонентов биосферы.
Динамическое единство биосферы возможно лишь в том случае, когда строительные и функциональные блоки универсальны. Если доказательства универсальности строительных блоков (аминокислоты, моносахариды, жирные кислоты и т. д.) были получены давно, то доказательства универсальности функциональных блоков появились лишь в последнее время.
Согласно концепции универсальных функциональных блоков, различные физиологические функции, выполняемые клетками разных тканей и органов, складываются из элементарных функций, реализуемых определенными функциональными блоками, число которых ограничено.
Функциональная специализация тканей обеспечивается благодаря различному сочетанию и количественному соотношению стандартных блоков, а изменение функциональных эффектов обусловлено их перераспределением.
Единство функциональных блоков у бионтов всех 5 царств обеспечивает как усвоение пищи (живое вещество предыдущего трофического звена), так и ресинтезы в следующем трофическом звене. Это обстоятельство делает понятным удивительное сходство важнейших характеристик пищеварительных ферментов, обеспечивающих процессы экзогенного питания, у организмов, находящихся на разных этапах филогенетического развития.
Еще более поразительным сходством характеризуются транспортные системы различных организмов, как простейших, так и высших позвоночных животных, обеспечивающие перенос через мембраны клеток молекулы аминокислот и сахаров. Поскольку биосфера существует благодаря круговороту вещества и потокам энергии, важную роль в которых играют гидролазы, разрушающие органические вещества продуцентов и консументов разного порядка, можно полагать, что жизнь на Земле сохраняется благодаря единству биосферы на уровне трофических взаимодействий в пределах биотических циклов.
Имеющиеся факты позволяют рассматривать единство механизмов экзотрофии не как результат стабилизирующего действия естественного отбора, направленного на поддержание определенного признака в определенном состоянии, а как результат сближения признаков у разных групп организмов. Эта конвергирующая функция позволяет сохранять существовавшее при формировании жизни единство и возможность взаимодействия различных звеньев трофических цепей между собой. Вышесказанное позволяет рассматривать биосферу как трофосферу, функционирующую по принципу трофостата, функции обратной связи в котором играют гидролазы, обеспечивающие деструкцию органического вещества.
Итак, благодаря важным открытиям в области физиологии, биохимии, цитологии и биологии в целом стала очевидной необходимость пересмотра ряда положений классической парадигмы, в частности теории сбалансированного питания.
Новая парадигма питания, не утрачивая точности физико-химических подходов, свойственных классической парадигме, обладает универсальностью, позволяющей анализировать все аспекты трофологии, за счет привлечения современных данных о неизвестных ранее механизмах начальных этапов ассимиляции пищи, о многообразии происходящих в пищеварительном тракте животных процессов, увеличивающих число потоков нутриентов и биологически активных веществ во внутреннюю среду организма, а также использования эволюционных и экологических подходов.
Как подчеркивалось ранее, рассмотрение в пределах одной науки микроскопических и планетарных процессов обусловлено единством механизмов экзотрофии в многоуровневой системе трофических связей. На одном полюсе этой системы стоит трофика как необходимое условие жизни, на другом — превращение и перемещение огромных масс веществ и энергии в биосфере в результате трофической иерархии и взаимосвязанности организмов в масштабах планеты. Это и позволяет воспринимать биосферу в определенном смысле как трофосферу, где пищевые связи образуют замкнутый контур. Грандиозность и различия масштабов, которыми оперирует трофология, не должны удивлять. Хотя носителями жизни служат отдельные организмы, в основе ее лежат элементарные процессы на молекулярном и клеточном уровнях, а в целом жизнь возможна лишь как планетарное явление. При этом на всех уровнях организации живых систем начальным звеном жизненного цикла является ассимиляция, служащая предметом трофологии.
6.2. ФИЗИОЛОГО-БИОХИМИЧЕСКИЕ ОСНОВЫ КОРМЛЕНИЯ РЫБ
|
|
|
|
Дата добавления: 2014-01-06; Просмотров: 702; Нарушение авторских прав?; Мы поможем в написании вашей работы!