КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Генетика психических заболеваний 4 страница
Таблица 4
Коэффициенты наследуемости скоростных характеристик двигательных реакций
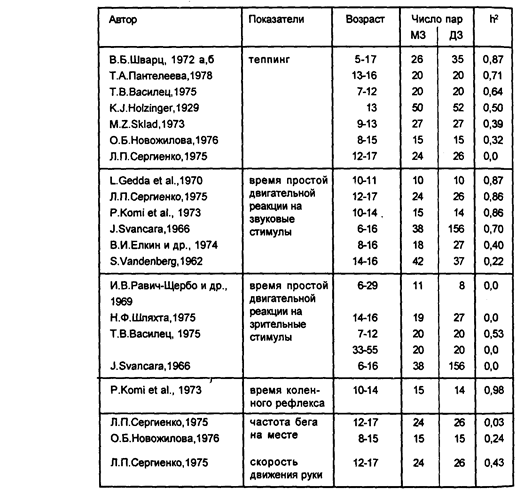
Большое количество фактов о наследственной обусловленности скоростных характеристик двигательных актов накоплено в рамках исследований свойств нервной системы близнецов, поскольку для диагностики таких свойств, как подвижность и лабильность, использовали различные двигательные методики (теппинг-тест, реакция выбора и т. д.).
С помощью двигательной методики изучалась подвижность нервной системы у близнецов (Канаев И.И., 1941; Василец Т.В., 1975, 1978 и др.), показатели которой оказались у МЗ близнецов очень похожими.
Н.Ф.Куркчи и Л.Н.Куркчи (1967) исследовали 3 пары ДЗ и 3 пары МЗ (8—17 лет) по методике А.Е.Хильченко. Испытуемым демонстрировалась движущаяся лента с изображениями круга, квадрата и треугольника, следующими в различном порядке. Испытуемый должен был на появление квадрата и круга нажимать кнопку соответственно правой и левой рукой. В качестве показателя подвижности использовалась максимальная скорость, при которой испытуемый не допускал больше 5% ошибок. Было обнаружено, что показатели подвижности у МЗ близнецов внутрипарно более сходны, чем у ДЗ близнецов.
В работах Т.В.Василец (1975, 1978) для определения подвижности были выбраны следующие методики: переделки сигнального значения раздражителей, измерения последействия стимулов, измерения темпа несложной деятельности. Все три экспериментальных приема были реализованы на базе методики Хильченко—Борисовой (двигательная реакция выбора из трех альтернатив при случайной последовательности стимулов и ускоряющемся темпе их предъявления). Во всех трех случаях использовались одни и те же раздражители, одна и та же двигательная реакция, и влияние силы нервной системы было элиминировано. Исследование проводилось с близнецами двух возрастных групп (7—11 и 33—55 лет), каждая возрастная группа включала по 20 пар МЗ и 20 пар однополых ДЗ близнецов. Была обнаружена разная степень генетических влияний на разные показатели. Наиболее отчетливую зависимость от генотипа продемонстрировал предельный темп реакции выбора. В изменениях латентных периодов влияния факторов наследственности не обнаруживается, а в динамике ошибок генетический контроль обнаруживается только в группе взрослых испытуемых.
В работе Т.А.Пантелеевой и Н.Ф.Шляхты (1978) исследовались некоторые показатели лабильности нервных процессов (куда входят и показатели теппинг-теста) у близнецов 13—16 лет (20 пар МЗ и 20 пар ДЗ). Обнаружено, что максимальный и оптимальный темп постукиваний зависит от генотипических факторов (оптимальный: у МЗ коэффициент корреляции равен 0,78, а ДЗ — 0,15; максимальный темп: у МЗ—0,69,уДЗ—0,25).
Генетический анализ времени простых сенсомоторных реакций дал противоречивые результаты (Gedda L. et al., 1970; Komi P. et al., 1973; Пантелеева Т.Д., 1975; Vandenberg S., 1962; Равич-Щербо И.В. с соавторами, 1965 и многие другие) — от явного влияния генотипа до отсутствия такового (табл. 4). Следует отметить, что в данных исследованиях сенсомоторная реакция рассматривалась как элементарный гомогенный акт, тогда как в современной психологии и физиологии эта реакция считается сложным по структуре актом, состоящим из независимых друг от друга компонентов — премоторного, связанного с восприятием и анализом поступающей стимуляции, и моторного, связанного с реализацией движения (Бойко Е.И., 1964). Соответственно и время реакции складывается из "премоторного" и "моторного" времени, то есть является суммарным показателем времени протекания, по меньшей мере, двух различных процессов. Поэтому возможно, что полученные расхождения в оценках наследственной обусловленности общего времени реакции объясняются различиями в использованных моделях двигательных реакций, акцентировавших одну или другую составляющую.
Действительно, генетический анализ временных характеристик отдельных компонентов общего времени реакции, проведенный Т.А.Пантелеевой (1977), выявил эти различия. В исследовании участвовали по 20 пар МЗ и ДЗ, у которых регистрировались простые двигательные реакции на звуки частотой 1000 и 500 Гц и интенсивностью 60 дБ. Кроме того, у всех испытуемых формировали двигательный навык реакции выбора из двух альтернатив (звуковые сигналы те же) с последующей ее переделкой. Анализировались три временных параметра двигательной реакции: 1) латентный период электромиограммы (ЭМГ); соответствующий премоторному времени; 2) время опережения ЭМГ-реакцией двигательного эффекта; 3) время самого движения. Сумма двух последних параметров дает моторное время. Эти параметры анализировались при формировании двигательного навыка на этапе врабатывания и на этапе автоматизации.
Оказалось, что генотипическая детерминированность временных характеристик двух компонентов "моторного" времени проявляется как в простой двигательной реакции, так и в реакции выбора на всех этапах ее формирования, автоматизации и переделки. Для "премоторного" времени аналогичные результаты получены только при простых произвольных реакциях и при автоматизированной реакции выбора. В период же "врабатывания", когда навык лишь формируется, зависимость "премоторного" времени от факторов генотипа не обнаруживается. Последним, судя по данным Т.А.Пантелеевой, и объясняется отсутствие генотипических влияний в суммарном времени реакции выбора на этапе "врабатывания". Таким образом, в исследовании выявилась зависимость генотип-средовых соотношений в вариативности временных параметров движения от уровня обученности: роль генетических факторов обнаруживается лишь на этапе сформированного навыка, когда двигательная реакция уже автоматизировалась и имеет стереотипный характер; на этапе же "врабатывания", когда в развернутой форме осуществляется осознанный контроль, большую роль играют факторы среды (Необходимо иметь в виду, что в некоторых сериях этого исследования получены значимые различия между МЗ и ДЗ по первым двум из упомянутых выше параметров, т. е. эти выборки не принадлежат одной генеральной совокупности и, строго говоря, не могут быть сопоставлены).
Анализ условий регистрации сенсомоторных реакций выявил также зависимость генотип-средовых соотношений в изменчивости временных характеристик двигательных реакции от модальности императивных стимулов. Психогенетические исследования двигательных реакций на зрительные стимулы свидетельствуют, за исключением проведенной на выборке близнецов подросткового возраста работы С. Ванденберга (Vandenberg S, 1962), о значительном влиянии генотипа на индивидуальные особенности временных характеристик. Вариативность же подобных показателей при использовании слуховых стимулов в большей мере определяется факторами среды. Особенно отчетливо это проявилось в работе И.Шванцары (Svancara J., 1971), регистрировавшего время простой двигательной реакции на слуховые и зрительные стимулы у одного и того же контингента испытуемых (таблица 4). Поскольку время реакции на стимулы разной модальности регистрировалось в стандартных условиях и биомеханическая структура движения была в обоих случаях одинакова, то различия в уровне генетического контроля могут объясняться, очевидно, только различиями в генетической детерминации процессов, протекающих в сенсорных системах. Это частично совпадает с данными Г.АПантелеевой (1977).
Наконец, в некоторых работах была установлена зависимость генотип-средовых соотношений не только от обу-ченности испытуемого и модальности стимула, но и от возраста испытуемых. В специальной серии экспериментов проведенной ТА.Пантелеевой (1977), изучалась генотипическая обусловленность времени простой двигательной реакции на стимулы разной интенсивности (звуковой сигнал частотой 1000 Гц и интенсивностью 40, 60, 80, 100 и 120 дБ) в разные периоды онтогенеза. В исследовании приняли участие дети 9—11 лет (по 20 пар МЗ и ДЗ, 48 человек-неродственников), 13—16 лет (также по 20 пар МЗ и ДЗ, 44 неродственника) и взрослые 33—56 лет (23 пары МЗ, 20 пар ДЗ, 40 неродственников). В младшей и старшей группах была зафиксирована зависимость индивидуальных особенностей времени реакции от наследственных детерминант, в подростковом возрасте эта зависимость не установлена. Интересно отметить, что в ряде исследований теппинга и времени реакции, свидетельствующих об отсутствии влияния генотипа на вариативность этих признаков, также принимали участие близнецы подросткового возраста (см. табл. 4). По-видимому, гормональные и психологические особенности подросткового возраста могут быть одной из причин неоднозначности получаемых результатов.
В работе К.Макнемара (McNemar Q., 1933) по исследованию тонкой координации было изучено 47 пар МЗ и 48 пар ДЗ близнецов старшего школьного возраста. Испытуемые выполняли двигательные пробы из Стэндфордской батареи моторных тестов: "Ротор" (Koerth W., 1927), "Тремометр" (Whipple G., 1910), "Вращение сверла", "Упаковка катушек", а также "Сортировка карт". Для всех тестов Р.Сишором (Seashore R., 1928) была выявлена хорошая ретестовая надежность: коэффициенты корреляции между первым и повторным (через 48 часов) тестированием достаточно высоки и колеблются от 0,75 до 0,94. К.Макнемаром было зафиксировано высокое внутрипарное сходство МЗ по показателям тестов: коэффициенты внутрипарной корреляции составили 0,95 для МЗ и 0,51 — для ДЗ ("Ротор"); 0,84 и 0,43 ("Тремометр"); 0,82 и 0,56 ("Вращение сверла"); 0,71 и 0,44 ("Упаковка катушек"); 0,84 и 0,39 ("Сортировка карт").
Большее внутрипарное сходство показателей МЗ было зафиксировано и в проведенном в те же годы исследовании А.Н.Миреновой (1934) при выполнении 4 парами МЗ и 6 парами ДЗ ряда двигательных проб (бросание и катание мяча в цель, прыжок в высоту с места). Выбор проб определялся предполагавшимися типами психомоторных процессов, одни из которых, по мнению А.Н.Миреновой, протекают автоматически и не требуют сложных регуляций (прыжки в высоту), другие — включают в себя сложную координацию движений в соответствии с определенной схемой (бросание мяча в цель — сложная координация относительно наглядного оптического поля; катание — координация по внутренней схеме). Средние внутрипарные разницы для МЗ и ДЗ были соответственно: по прыжкам в высоту— 1,75 и 7,6 см; по бросанию мяча в цель — 0,25 и 1,47 очка: при катании мяча в цель — 15 и 23,8 см. Таким образом, различия МЗ близнецов по первому тесту в 4 раза, по второму — в 6 раз, по третьему — в 1,6 раза меньше, чем ДЗ, то есть, по мнению А.Н.Миреновой, более элементарные по своей координационной структуре двигательные действия в большей мере контролируются генотипом.
В проведенном почти через 30 лет исследовании С.Ванденберга (Vandenberg S., 1962) наряду с моторными тестами из Стэндфордской батареи использовались тесты "Лабиринт", "Рисунок с зеркала", а также методика "Ловкость обращения с пинцетом". При определении двигательных способностей 42 пар МЗ и 37 пар ДЗ были получены достаточно высокие коэффициенты наследуемое™ (от 0,52 до 0,71) результатов выполнения практически всех проб, кроме тестов "Лабиринт" и "Тремометр"; отмечено, что точность движений кисти рук в большей степени детерминируется генотипом, чем более грубые движения всей руки.
Исследования К.Макнемара и Т.А.Пантелеевой интересны тем, что в них специально прослеживалось влияние обучения на генотипическую обусловленность индивидуальных особенностей координации. В общей форме вопрос здесь ставится следующим образом: если в результате тренировки различия между МЗ и ДЗ по уровню внутрипарного сходства меняются, то это означает, что приобретаемый индивидуальный опыт может менять генотип-средовое соотношение в изменчивости данного признака.
Для проверки этого предположения К.Макнемар провел специальную серию экспериментов с обучением: тест "Ротор" испытуемый выполнял 70 раз, "Упаковку катушек" и "Сортировку карт" — по 28 раз. Успешность выполнения теста оценивалась в баллах, и, кроме того, регистрировалось время выполнения каждого теста. Серия с обучением состояла из семи частей — по 10 упражнений в каждой для теста "Ротор" и по 4 — для двух других. Эффект обучения был выявлен во всех тестах (см. таблицу 5).
Таблица 5
Коэффициенты внутриклассовой корреляции МЗ и ДЗ близнецов в начале и в конце тренировки (по Макнемару К., 1933)
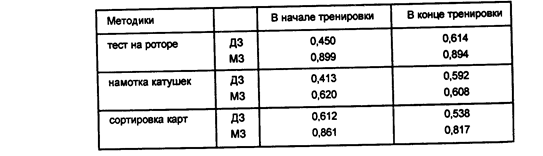
Как видно из таблицы 5, сходство показателей МЗ близнецов по успешности выполнения тестов "Ротор" и "Упаковка катушек" не зависит от обучения, хотя сходство ДЗ по мере обучения увеличивалось. В выполнении "Сортировки карт" в обеих группах наблюдалось незначительное уменьшение внутрипарного сходства. Различия между начальным и конечным коэффициентом корреляции у ДЗ равнялись: по тесту "Ротор"— 0,157±0,081 и по "Упаковке катушек"— 0,174±0,079; эти же коэффициенты ДЗ для временных показателей выполнения увеличились по тесту "Ротор" с 0,36 до 0,54 и по "Упаковке катушек" — с 0,36 до 0,58. Подсчет коэффициентов внутрипарной корреляции по всём частям серии выявил увеличение сходства ДЗ близнецов уже на начальной стадии обучения.
Осуществленный Т.А.Пантелеевой (1977) анализ индивидуальных особенностей временных характеристик двигательной реакции выбора на разных этапах формирования навыка также свидетельствует об уменьшении генетических влияний на этапе "врабатывания", что связано, как считает Т.А.Пантелеева, с высокой степенью осознанности и произвольной регуляцией движения.
В целом в исследованиях двигательных особенностей человека, оцениваемых по поведенческим параметрам выполнения различных моторных задач, было установлено весьма значительное влияние факторов генотипа в межиндивидуальной вариативности показателей гибкости, относительной мышечной силы, скоростно-силовых характеристик выполнения спортивных упражнений и временных характеристик коленного рефлекса. Индивидуальные же особенности ловкости, абсолютной мышечной силы, напротив, формируются в основном под влиянием факторов среды. Результаты исследований теппинга, сенсомоторных реакций и особенностей тонкой координации оказываются неоднозначными, что, вероятно, объясняется их сложной структурой, включающей разнородные и независимые друг от друга компоненты, и особенностями их регуляции. Кроме того, как показали некоторые исследования, влияние генетических факторов на межиндивидуальную вариативность двигательных качеств может существенно меняться в онтогенезе.
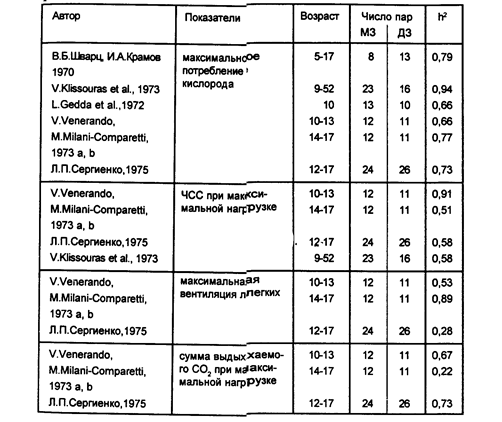
3. В генетических исследованиях физиологических факторов реализации движений в основном изучались спортивные способности. Первыми работами этого направления стали исследования В.Б.Шварца и И.А.Крамова (1970), а также В.Клиссаураса (Klissouras V., 1971), установивших значительное влияние генетических факторов на межиндивидуальную вариативность некоторых параметров кровообращения и максимального потребления кислорода (МПК) при выполнении спортивных движений. Исследования МПК оказываются наиболее интересными. Этот показатель (от которого в решающей мере зависит успешность в циклических видах спорта) приблизительно одинаков в разных этнических группах, не меняется в онтогенезе, мало тренируем и оказывается высоко генетически детерминированным (см. В.Б.Шварц, 1978). Если учесть, что спортсмены международного класса имеют показатели МПК, значительно превышающие их среднепопуляционную величину, то, по-видимому, индивидуальный уровень МПК может служить информативным признаком при прогнозировании спортивной успешности.
Межиндивидуальная вариативность показателей частоты сердечных сокращений (ЧСС) при максимальной нагрузке и сумма выдыхаемого СО, при максимальной работе также зависят от наследственных факторов, тогда как индивидуальные особенности ЧСС в покое и максимальная вентиляция легких зависят, как правило, от факторов среды (таблица 6).
Таблица 6
Коэффициенты наследуемости некоторых физиологических показателей
4. Использование в генетике поведения метода регистрации потенциалов мозга, связанных с движением (ПМСД), дает возможность оценить вклад генетических факторов в изменчивость биоэлектрических коррелятов индивидуальных особенностей организации и построения движений.
Использование процедуры усреднения дало возможность исследователям выделять в записи ЭЭГ изменения потенциала, связанные с организацией и выполнением, двигательного акта (отрезки ЭЭГ суммируются до, во время и после двигательной реакции). В комплексе потенциалов (потенциал готовности, компоненты PI, N2, Р2, N3 и РЗ), предшествующих и сопровождающих реализацию двигательного акта, находят свое отражение процессы подготовки, реализации и оценки движения.
В исследовании С.Б.Малых (1986, 1988) было показано, что в ситуации простого нажатия на кнопку во всех зонах регистрации, кроме левой лобной области, уровень внутирипарного сходства амплитудно-временных показателей практически всех компонентов ПМСД выше у МЗ, чем у ДЗ. В левой лобной области отмечается противоположная тенденция: внутрипарное сходство амплитуд и временных показателей всех компонентов, кроме реафферентного потенциала Р2, у ДЗ — даже несколько выше. В ситуации прогноза различия между МЗ и ДЗ в уровне внутрипарного сходства как временных, так и амплитудных параметров ПМСД, УНВ и РЗОО увеличивались, причем в ситуации неоднозначного прогноза эти тенденции становились отчетливее. При прогнозе равновероятных событий сходство амплитуд практически всех компонентов потенциала, кроме ПГ и РЗ левой лобной области и ПГ, N3, РЗ правой центральной области, было значимо выше у МЗ. Сходство временных параметров в целом намного ниже и у МЗ, и у ДЗ. При заведомо истинном прогнозе наблюдалась аналогичная картина, хотя различия МЗ и ДЗ по амплитудно-временным параметрам ПМСД, УНВ и Р 300 в этой экспериментальной ситуации ниже, чем при прогнозе равновероятных сигналов. В целом, чаще всего влияние генотипа обнаруживалось в ситуации прогноза равновероятных событий (54% всех анализируемых показателей), значительно реже — при простых произвольных движениях (20%). Ситуация заведомо истинного прогноза занимала промежуточное положение (46%). Таким образом, эти результаты свидетельствуют о зависимости соотношения генетических и средовых факторов в вариативности ПМСД от места движения в общей структуре двигательного акта.
Анализ влияния генетических факторов на индивидуальные различия ПМСД в ходе развития показал, что высокие и значимые корреляции у МЗ близнецов наблюдаются в основном для последвигательных компонентов ПМСД. Данные о генетическом контроле ПМСД в ходе развития свидетельствуют об изменении соотношения генетических и средовых факторов с возрастом, что, возможно, связано со сменой мозговых механизмов организации двигательного действия в онтогенезе (Кузнецова И.В., 1992; Малых С.Б., Кузнецова И.В., 1992; Malykh S., Orekhova E., 1995; Орехова Е.В., 1996).
В целом нейрофизиологическое исследование двигательного акта показало, что в детерминации изменчивости амплитудно-временных показателей компонентов ПМСД, отражающих особенности центральных процессов организации и построения двигательных актов, наблюдаются существенные различия в зависимости от места движения в структуре действия и от уровня нейрофизиологического обеспечения.
Приложение
к лабораторному занятию № 11-12 «Генетическая психофизиология»
1. В последние годы на стыке психогенетики и психофизиологии возникло новое направление исследований — генетическая психофизиология. Основное внимание специалисты в области психогенетики уделяли исследованию наследственной детерминации когнитивной и личностной сферы человека, тогда как более простые психологические и психофизиологические характеристики оставались практически не исследованными. Это весьма существенный пробел, поскольку особенности структуры и функций нервной системы являются опосредующим звеном в цепи "ген — поведение". Между генами индивида и его поведением находится целый ряд уровней организации: клеточный (физико-химические процессы внутри клетки), межклеточный (физико-химические процессы между клетками), уровень целостных органов (их структура и функции) и уровень целостной индивидуальности. Для того чтобы понять механизмы влияния генотипа на поведение, необходимо исследовать генетическую изменчивость структуры и функций нервной системы на всех уровнях — от биохимических процессов внутри клетки до суммарных биоэлектрических процессов целого мозга (рисунок I) (Фогель Ф., Мотульски А., 1990).
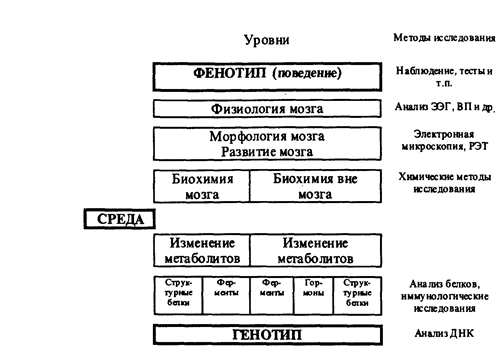
Рисунок I
Схема уровней исследования генетической изменчивости
функций мозга (по Фогелю Ф., Мотульски А., 1990)
Конечной целью генетического анализа является определение местонахождения гена и типа наследования, детерминирующего тот или иной признак, тем не менее, первый этап генетического анализа всегда направлен на установление самого факта наследственной детерминации психофизиологических показателей. Ведь сначала необходимо выяснить, имеют ли гены отношение к формированию индивидуальных различий в психофизиологических характеристиках. Решению именно этой проблемы посвящены сегодня все усилия исследователей в области генетической психофизиологии.
|
|
Дата добавления: 2014-11-08; Просмотров: 887; Нарушение авторских прав?; Мы поможем в написании вашей работы!