КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Исследования генетической детерминации ЭЭГ человека
|
|
|
|
ЭЭГ состояния покоя
Генетические исследования ЭЭГ начались в 30-е годы, то есть вскоре после сообщения Бергера о регистрации ЭЭГ у человека. В первых исследованиях ЭЭГ преобладали методы визуального анализа. В них ЭЭГ человека фактически рассматривалась как некий паттерн, то есть характерный рисунок. При таком подходе ЭЭГ выступала как качественный признак. Исследователи, регистрируя ЭЭГ у родственников (чаще всего у близнецов), визуально выделяли определенные типы кривых и пытались их классифицировать. В результате родственники могли оцениваться как конкордантные или дискордантные по форме ЭЭГ.
Первые же исследования показали высокое сходство общего рисунка ЭЭГ у членов МЗ пар. Как правило, различия между МЗ близнецами не превышали различий между двумя отрезками ЭЭГ одного человека, записанными в разные дни (Davis H., Davis P., 1936; Raney E., 1939; Lennox W. et al., 1945). Однако наиболее интересными можно считать данные, полученные Джуель-Нильсеном и Харвальдом (Juel-Nielsen N., Harvald В., 1958), которые показали, что различные параметры ЭЭГ даже у разлученных МЗ близнецов практически идентичны. Исследования ЭЭГ у разлученных близнецов проводились и в последующие годы (Lykken D., 1982; Boushard Т., 1990), на более солидных выборках и с применением более современных методов анализа. Все полученные результаты указывают на высокий уровень сходства параметров ЭЭГ у разлученных близнецов. Поскольку у разлученных МЗ близнецов нет никаких общих средовых факторов, то все их сходство возникает только за счет генетической идентичности. Соответственно, данные о поразительном подобии ЭЭГ у разлученных близнецов говорят о том, что в основе семейного сходства лежат не средовые, а генетические причины.
|
|
|
Паттерны ЭЭГ у монозиготных близнецов оказались настолько похожими, что этот признак предлагалось использовать при диагностике зиготности близнецов наряду с цветом глаз, волос и другими морфологическими особенностями.
Анализ ЭЭГ как целостного паттерна используется в генетических работах и в наши дни. Наиболее полно он представлен в исследованиях ЭЭГ, ведущихся под руководством Ф.Фогеля в Германии (Fogel F. et al., 1979,1981). Некоторые из паттернов ЭЭГ, для которых Ф.Фогелем были установлены простые (менделевские) типы наследования, внесены в каталог "Наследственные признаки человека" (Маккьюсик В., 1976). Однако индивидов с такими характерными паттернами ЭЭГ в популяции встречается немного (не более 5%). Их можно считать крайним выражением всего разнообразия, наблюдаемого в популяции. Примером таких "крайних" типов являются выделенные Ф.Фогелем паттерны, получившие название "монотонные альфа-волны" и "низковольтная ЭЭГ" (рисунок 3). Для них установлен аутосомно-доминантный тип наследования. Пробанды, обладающие этими типами ЭЭГ, отличаются некоторыми психологическими и биохимическими характеристиками (Fogel F. et al., 1979; Fogel F., Propping P., 1981).
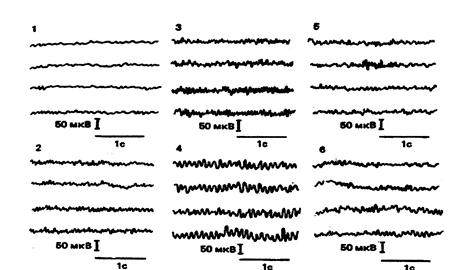
1. Низковольтная
2. Низковольтная пограничная (бета-волны)
3. Затылочные медленные бета-волны
4. Монотонные альфа-волны
5. Фронто-прецентральные бета-волны
6. Диффузные бета-волны
Рисунок 3
Наследуемые типы ЭЭГ по Ф. Фогелю
Более распространен, однако, подход к ЭЭГ как к количественному признаку, поскольку все ритмические составляющие ЭЭГ могут быть охарактеризованы всевозможными количественными измерениями (частота, амплитуда, спектральная плотность, когерентность и т.д.). Такой подход следует считать более перспективным, так как он охватывает всю изменчивость, представленную в популяции.
Количественные показатели также фигурируют в работах Ф.Фогеля (Vogel F., 1958, 1962а, 19626). Первое же его исследование было проведено на большой выборке близнецов (100 пар МЗ и 98 пар ДЗ). Регистрация ЭЭГ у близнецов велась не только в состоянии покоя, но и при различных функциональных нагрузках (гипервентиляция, гипоксия и сон). Фиксировался ряд показателей: доминирующая частота ЭЭГ; альфа-индекс (процент времени, в течение которого в ЭЭГ наблюдается выраженный альфа-ритм); амплитуда альфа-ритма; процент совпадения альфа-волн по фазе; регулярность альфа-ритма. Анализ количественных параметров ЭЭГ показал, что МЗ близнецы практически идентичны по показателям альфа-индекса ЭЭГ, амплитуды и частоты альфа-ритма. Различия между МЗ близнецами были не больше, чем у одного и того же человека при повторной регистрации ЭЭГ. Это дало Ф.Фогелю основание для вывода о высокой наследуемости ЭЭГ.
|
|
|
Последующие работы подтвердили выводы Фогеля. В большинстве исследований, несмотря на разные размеры выборок (часто небольшие) и несовпадение методов регистрации и анализа, монотонно обнаруживалось высокое внутрипарное сходство МЗ близнецов по любым характеристикам альфа-ритма (Inouye E., 1961; Kamitake M., 1963; Young J., 1972; Мешкова Т.А., 1976; Анохин А.П., 1987 и др.).
С развитием методов спектрального анализа ЭЭГ и в генетических работах все чаще стали фигурировать более сложные, нежели частота, амплитуда, альфа-индекс, количественные характеристики ЭЭГ. Среди них наиболее распространенными количественными оценками являются суммарные энергии и спектральные плотности отдельных ритмов. [Суммарные энергии и спектральные плотности являются параметрами, которые различным образом отражают представленность данного ритма в ЭЭГ. Суммарная энергия частотного диапазона ЭЭГ представляет собой абсолютную амплитуду ритма данного диапазона. Спектральные плотности мощности спектров отдельных диапазонов представляют собой величины, нормированные относительно мощности спектра всего диапазона частот]
Спектральный анализ ЭЭГ использовался в работе Д. Ликкена с соавторами (Lykken D. et al., 1974). Спектральные характеристики ЭЭГ оказались чрезвычайно похожими у МЗ близнецов. Их внутрипарные корреляции были сопоставимы с таковыми для морфологических признаков — длины и веса тела. В зависимости от частотного диапазона величина корреляций колебалась в пределах от 0,76 до 0,86. В то же время в группе ДЗ корреляции оказались очень низкими — от -0,20 до 0,15. Это значительно ниже, чем можно ожидать, исходя из предположения об аддитивности генетических влияний. Правда, условия регистрации ЭЭГ в исследовании Ликкена несколько отличались от обычных: ЭЭГ записывалась у испытуемых в состоянии гипноза. Повторный эксперимент, проведенный в 1982 году (Lykken D. et al., 1982), дал те же результаты. Кмз в несколько раз превышал г, а г во многих случаях был близок к нулю, что свидетельствует о неаддитивном типе наследования спектральных плотностей ритмических составляющих ЭЭГ. Д.Ликкен (Lykken D., 1982) называет такие признаки, по которым сходство МЗ значительно превышает сходство ДЗ, эмерджентными (emergenic traits). Наследственная передача таких признаков обеспечивается полигенными механизмами. Для проявления определенного признака необходимо определенное сочетание нескольких генов, передающихся от родителя к потомку случайным образом. Поскольку МЗ близнецы имеют идентичный набор генов, их сходство по эмерджентным признакам также достаточно велико. Для ДЗ близнецов и других близких родственников вероятность наличия одинакового набора генов убывает в геометрической прогрессии в соответствии с увеличением числа генов, определяющих данный конкретный признак.
|
|
|
Первые отечественные генетические исследования ЭЭГ были начаты в 70-е годы в русле исследования свойств нервной системы коллективом сотрудников под руководством Б.М.Теплова и В.Д.Небылицына, а затем И.В.Равич-Щербо. Это работы Н.Ф.Шляхты, Г.А.Шибаровской, Т.А.Мешковой, Н.В.Гавриш, С.Б.Малых. Во всех этих исследованиях, проводившихся на близнецах, в целом по большинству количественных параметров ЭЭГ отмечается более высокое внутрипарное сходство МЗ близнецов по сравнению с ДЗ близнецами.
Используя широкий набор количественных параметров ЭЭГ, полученных в результате ручной и автоматической обработки, Т.А.Мешкова (1978) провела сравнение оценок наследуемости ЭЭГ различных областей мозга — лобных, центральных, теменных, затылочных и височных — в симметричных отведениях с обоих полушарий. Оказалось, что более высокие оценки наследуемости имеют показатели альфа-ритма, по сравнению с другими частотными диапазонами, и ЭЭГ правого полушария, по сравнению с левым. Доля генетических компонент фенотипической дисперсии колебалась от 46% до 96% для разных параметров и разных областей мозга. Интересно, что наименьшие генотипические влияния практически по всем параметрам были характерны для ЭЭГ левой височной области коры. По мнению автора, это связано с филогенетическим возрастом этой области мозга (она появляется достаточно поздно в филогенезе), длительностью ее созревания в онтогенезе, а также особой ролью в осуществлении речевых функций.
|
|
|
Исследование Н.В.Гавриш и С.Б.Малых (1994), проведенное на близнецах 6—8 лет (26 пар МЗ и 22 пары ДЗ), в котором выделялось три (по АлферовойВ.В., Фарбер Д.А., 1990) основных поддиапазона альфа-ритма, позволило сделать заключение о различной природе межиндивидуальной изменчивости отдельных частотных компонентов альфа-ритма. При этом наблюдается следующая закономерность: чем позже в онтогенезе соответствующий компонент альфа-ритма становится доминирующей формой активности, тем меньшую роль в семейном сходстве играют генетические факторы. Один из выводов, сделанных авторами, заключается в том, что при генетическом анализе ЭЭГ неправомерно сопоставлять данные разных возрастных групп по параметрам только широкополосного альфа-ритма.
Структура фенотипической изменчивости некоторых количественных параметров ЭЭГ анализировалась также в семьях туркменской популяции (Анохин А.П.,1987). Для большинства показателей абсолютной мощности спектров вклад генетических факторов оказался достаточно высоким, за исключением бета-диапазона в затылочных отведениях. Аддитивная генетическая компонента изменчивости (GА) варьировала от 0% для бета-диапазона в затылочной области до 80% для бета-диапазона в лобных отведениях. Как правило, фенотипическое сходство между сибсами превышало сходство между родителями и детьми. Это объясняется как эффектами доминирования, так и влиянием систематических общесемейных средовых факторов. При анализе относительных мощностей частотных диапазонов было выявлено две группы показателей: 1) показатели с низкими оценками наследуемости — характеристики электрической активности лобных областей мозга и тета-активность височных областей; 2) показатели с высоким уровнем наследуемости — характеристики ЭЭГ височной и затылочной областей.
К сожалению, в большинстве генетических исследований ЭЭГ либо обследовались небольшие контингента испытуемых одного возраста, либо возрастной диапазон был слишком велик, что для популяционных исследований является большим недостатком. От размера выборки зависит точность оценки компонент дисперсии исследуемых признаков.
Возможность оценки значения того или иного эффекта зависит от целого ряда факторов: действительной величины эффекта в популяции (чем больше эффект, тем легче он выявляется); уровня достоверности, выбранного в качестве критерия для отвержения нулевой гипотезы об отсутствии влияния эффекта; размера выборки (чем больше размер выборки, тем более тонкие эффекты мы можем оценить). Исследования, в которых обследуются большие выборки испытуемых, дают возможность более точной оценки всех эффектов, особенно это касается тех случаев, когда величина эффекта в популяции невелика.
В этом плане своеобразной вехой являются опубликованные недавно результаты исследования, проведенного голландскими учеными из Свободного университета г.Амстердама (van Beijsterveldt С. et al.,1996; van Baal С. et al., 1996). Остановимся на нем более подробно, поскольку оно, в определенном смысле, является образцовым (как по выборке испытуемых, так и по использованным методам анализа). Во-первых, следует отметить, что для определения зиготности близнецов использовались не только опросники и биохимия крови, но и "генная дактилоскопия" (Генная дактилоскопия (DNA fingerprint) — способ установления генетической идентичности, основанный на анализе спектров образцов ДНК. Уникальность каждого индивида определяется при этом на уровне последовательности нуклеотидов. Идентичность спектров ДНК у близнецов однозначно указывает на их монозиготность, а различия между спектрами говорят о дизиготности близнецов.), что обеспечивает 100-процентную надежность определения зиготности.
Во-вторых, впервые близнецовое исследование ЭЭГ проведено на столь представительной выборке: 71 пара МЗ и 96 пар ДЗ близнецов пяти лет в исследовании К. ван Баал и др. (1996) и 91 пара МЗ и 122 пары ДЗ близнецов 16 лет в исследовании К. ван Бейстервельдт и др. (1996). В-третьих, регистрация ЭЭГ осуществлялась в 14 отведениях. В-четвертых, помимо анализа каждого отведения в отдельности, авторы рассмотрели взаимосвязи между биоэлектрической активностью различных областей коры (показатели когерентности). В-пятых, для оценки генетических и средовых компонент фенотипической дисперсии использовали новейшие методы генетического анализа, в частности, метод структурного моделирования.
Сначала подробно остановимся на анализе ЭЭГ отдельных отведении. Рассмотрим вначале результаты, полученные на выборке пятилетних близнецов. ЭЭГ детей записывали в состоянии покоя (3 минуты с открытыми глазами и столько же с закрытыми). Анализировали спектры мощности дельта(1,5—3,5 Гц), тета (4—7,5 Гц), альфа (8—12,5 Гц), и бета (13—25 Гц) диапазонов.
В пятилетнем возрасте у детей доминируют медленные волны (дельта, тета и альфа-1 в сумме составляют почти 92% общей вариативности сигнала), тогда как более быстрые волны (альфа-2, бета-1 и бета-2) представлены меньше (лишь 8%). Наибольшая мощность спектра характерна для тета-активности. Результаты свидетельствуют о высокой наследуемости тета, альфа-1 и альфа-2 частот: в среднем оценки наследуемости для показателей абсолютной мощности тета-диапазона равны 81%, для альфа-1 и альфа-2 диапазонов — 81% и 78% соответственно. Для других диапазонов влияние генетических факторов несколько меньше, но тем не менее наследственные факторы объясняют большую часть наблюдаемых индивидуальных различий в показателях дельта- и бета-диапазонов (средние значения наследуемости составляют 55% для дельта диапазона, 73% — для бета-1 и 64%— для бета-2). Генетические влияния на показатели относительных мощностей дельта- и бета-диапазонов несколько больше, чем для абсолютных спектров (значения коэффициента наследуемости составляют 63% для дельта-диапазона; 76%— для тета-дипазона; 71%— для альфа-1 и 72% — для альфа-2 диапазона; 68% и 65% — для обоих бета-диапазонов).
Преимуществом данного исследования является и то, что в нем принимали участие также и разнополые близнецы, что позволило оценить половые различия в наследуемости характеристик ЭЭГ. Корреляции разнополых и однополых ДЗ близнецов оказались примерно равными, что свидетельствует об одинаковом уровне генетических и средовых влияний в группах мальчиков и девочек. Метод подбора моделей, использованный для генетического анализа, показал, что полученные данные лучше всего описываются простой генетической моделью (АЕ).
При исследовании 16-летних близнецов использовались те же отведения и методы анализа, что и на выборке пятилетних детей. В этом возрасте, так же как и в 5 лет, практически для всех показателей наблюдалось высокое сходство МЗ близнецов (rмз около 0,85); корреляции ДЗ были примерно вполовину меньше. Использование метода подбора моделей показало, что для большинства ритмов во всех областях мозга простая генетическая модель наилучшим образом описывает полученные данные.
Рассмотрим результаты одномерного генетического анализа.
Дельта-диапазон. Корреляции МЗ близнецов в этой частотной области были ниже, чем в других. Корреляции ДЗ близнецов в среднем были в два раза меньше, чем МЗ. Подбор моделей показал, что наилучшие результаты дает простая генетическая модель. Наследуемость ЭЭГ-характеристик дельта-диапазона во фронтальных областях составляла около 70%, тогда как для задних областей она была выше (80%).
Тета-диапазон. Для тета-диапазона внутрипарные корреляции МЗ оказались выше (в среднем для всех отведении около 0,9); корреляции ДЗ в среднем составляли около половины корреляций МЗ. Уровень половых различий в генетических влияниях был невелик. Как и для дельта-диапазона, простая генетическая модель наилучшим образом описывает полученные данные, за исключением отведении СЗ и РЗ. Для этих отведении данным лучше всего соответствует генотип-средовая модель, учитывающая половые различия.
Альфа-диапазон. Данные по мощности альфа-ритма лучше всего описываются простой генетической моделью. Коэффициент наследуемости (усредненный по всем отведениям) составляет около 89%. Половых различий в генетических влияниях не обнаружено.
Бета-диапазон. Корреляции ДЗ близнецов-девочек оказались несколько ниже, чем можно было предположить, исходя из степени родства. Корреляции МЗ были на том же уровне, что и для альфа-диапазона. Подобные результаты указывают на наличие генетических эффектов, обусловленных доминированием. Результаты подбора моделей показали, что для двух отведении (РЗ и Т5) наилучшим образом подходит аддитивно-доминантная модель, а для оставшихся областей — простая генетическая модель.
Многомерный анализ ЭЭГ. Многомерный анализ был проведен только для показателей альфа-диапазона, поскольку он является доминирующей частотой в ЭЭГ. Этот способ анализа позволяет оценить, в какой степени одни и те же гены вносят свой вклад в наблюдаемую дисперсию ЭЭГ различных зон мозга. Наличие высоких генетических корреляций между всеми отведениями подтверждает предположение о том, что одни и те же генетические воздействия лежат в основе ЭЭГ разных отведении.
Результаты двумерного анализа свидетельствуют о том, что ковариации между симметричными отведениями правого и левого полушарий обусловлены, как правило, одним общим генетическим фактором, за единственным исключением: ковариация между отведениями Fpl—Fp2 обусловлена также и специфическим генетическим фактором.
Ковариация между симметричными отведениями обусловлена в основном генетическими факторами (вклад генетических факторов составляет почти 90%), хотя свой вклад вносит и фактор различающейся среды.
В целом результаты работы голландских исследователей свидетельствуют о значительном влиянии генетических факторов на индивидуальные различия в спектральных характеристиках ЭЭГ, которое увеличивается с возрастом.
Теперь подробно рассмотрим результаты голландского исследования, касающиеся анализа когерентности ЭЭГ.
Сначала несколько слов о самой когерентности. Показатели когерентности обычно используют для оценки синхронности ЭЭГ в двух различных отведениях.
Когерентность g 2xy (w) является оценкой корреляции процессов x(t) u y(t) для каждой частотной составляющей этих процессов в области определения их спектров. Близкая к 1 когерентность на частоте w для фазы Фxy (w), близкой к нулю, соответствует высокой положительной корреляции в обычной статистике; для фазы, близкой к 1800 — высокой отрицательной корреляции. Функция полностью независимых процессов равна нулю. Одновременное определение когерентности и фазы позволяет наиболее полно изучать корреляционные свойства процессов.
Тэтчер (Tatcher R., 1987) считает, что когерентность между двумя отведениями отражает наличие аксональных взаимосвязей между соответствующими областями мозга. Число и мощность связей находят свое отражение в показателях когерентности. Высокая когерентность указывает на то, что ЭЭГ различных зон мозга синхронизированы, и рассматривается как показатель "связанности" областей мозга, поскольку синхронизированные колебания в нейронных сетях наблюдаются только в том случае, если они связаны между собой.
Дело в том, что отдельные области мозга не работают изолированно друг от друга, их взаимодействие координируется сложными нейронными цепями, которые связывают их между собой. Число этих кортико-кортикальных связей у человека очень велико, и их развитие продолжается вплоть до 20-ти лет. Предполагается (Changeux J., Danchin A., 1976), что общая программа формирования кортико-кортикальных связей находится под генетическим контролем, но общее число синапсов слишком велико для того, чтобы использовалась какая-либо специфическая генетическая программа. Действительно, морфология мозолистого тела у МЗ близнецов более сходна, чем у неродственников, что свидетельствует в пользу влияния генотипа. С другой стороны, полной морфологической идентичности у членов МЗ пар нет, что говорит о возможном участии среды в формировании морфологических особенностей центральной нервной системы. Вероятно, только общий план нейронных взаимосвязей генетически запрограммирован (Huttenlocher P., 1994), тогда как более тонкая настройка формируется во взаимодействии со средой (Wiesel Т., 1994).
Развитие центральной нервной системы — эпигенетический процесс. ЦНС развивается во взаимодействии со средой, и результатом этого развития является модификация как ЦНС, так и среды (Benno R.,1990; Molenaar P. et al., 1993).Таким образом, при анализе индивидуальных различий во взаимосвязях между областями мозга, наряду с генетическими факторами, можно ожидать влияния специфического индивидуального опыта.
Следует отметить, что когерентность изменяется в ходе созревания мозга. Исследование Р.Тэтчер (Tatcher R. et al., 1987, 1994), проведенное методом поперечных срезов на большой выборке испытуемых разного возраста — от младенцев до взрослых, показало большие изменения когерентности в онтогенезе.
Исследования К. ван Бейстервельдт и К. ван Баал (van Beijsterveldt С., 1996; van Baal С., 1997) являются однимииз первых работ, посвященных оценке вклада генетических факторов в индивидуальные различия во взаимосвязях между различными областями мозга.
Исследование К. ван Баал (1997) выполнено на близнецах 5 лет (167 пар). Когерентность подсчитывалась в тета-диапазоне (4,0—7,5 Гц) между префронтальными, фронтальными, центральными, париетальными и затылочными областями.
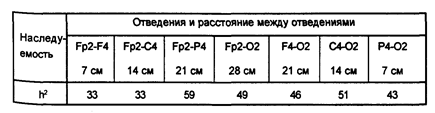
Одномерный генетический анализ выявил достаточно сильные генетические влияния для всех показателей когерентности. Корреляции в группе МЗ близнецов всегда были выше, чем в группе ДЗ, величина оценок наследуемости когерентности колебалась от 30% до 71% (таблицы 1 и 2)
Таблица 1
Оценки наследуемости (в широком смысле) когерентности в тета-диапазоне в левом полушарии у детей 5 лет

Таблица 2
Оценки наследуемости (в широком смысле) когерентности в тета-диапазоне в правом полушарии у детей 5 лет
Анализ когерентности ЭЭГ у близнецов юношеского возраста (16 лет) (van Beijsterveldt С., 1996) также выявил наличие генетических влияний в индивидуальных различиях по этому показателю. Когерентность вычислялась во всех частотных диапазонах для тех же самых отведении, что и в предыдущей работе. Выборка включала в себя 89 пар МЗ, 72 пары однополых ДЗ и 46 пар разнополых ДЗ.
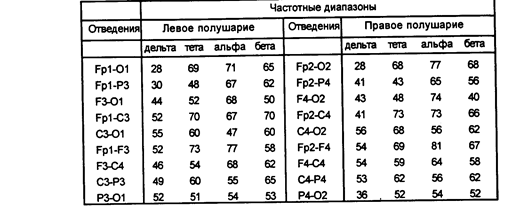
Результаты свидетельствуют о значительном влиянии генетических факторов (не менее 50%) в индивидуальных различиях когерентности во всех анализируемых частотных диапазонах. Наибольшая доля вариативности, обусловленная генетическими причинами, наблюдается для "быстрых" частотных диапазонов. Показатели наследуемости (усредненные по всем отведениям) составляют 60%, 65% и 60% для тета-, альфа- и бета-диапазонов соответственно. Наследуемость когерентности для дельта-диапазона ниже.
Межполушарных различий в наследуемости когерентности не было обнаружено (таблица 3).
Таблица 3
Коэффициенты наследуемости когерентности у близнецов 16 лет
Поскольку, как мы уже упоминали, когерентность может служить индикатором того, в какой степени различные области мозга связаны между собой, полученные результаты свидетельствуют о значительном влиянии генетических факторов на изменчивость связей между областями мозга 16-летних детей.
Как отмечают авторы работы, наследуемость когерентности несколько ниже, чем наследуемость показателей мощности спектра, для которых усредненный по всем частотным диапазонам и отведениям коэффициент наследуемости составляет 85%. Меньшая наследуемость когерентности, вероятно, обусловлена более низкой стабильностью этой характеристики. Очевидно, что увеличение ошибки измерения, которая входит в общую дисперсию, приводит к уменьшению наследуемости. В ряде работ оценивалась ретестовая надежность показателей когерентности (Gasser Т. et al., 1987; Dunkin J. et al., 1994; Harmony Т. et al., 1993). Исследования были проведены на различных по размеру выборках испытуемых разного возраста, с различным интервалом между повторными измерениями. Гессер (Gasser Т. et al., 1987) обнаружил, что при интервале между измерениями в один год для молодых испытуемых характерны более низкие ретестовые корреляции (0,40), чем для пожилых (0,60). Таким образом, было установлено, что воспроизводимость показателей когерентности ниже, в сравнении с показателями мощности спектра. Соответственно, меньшая надежность когерентности может быть причиной более низких оценок наследуемости.
3. КГР в состоянии покоя. Все немногочисленные работы, посвященные анализу КГР в состоянии покоя, были проведены с использованием близнецового метода. Первое известное генетическое исследование электрической активности кожи было проведено в 1934 году Карменом (цит. по Lader M., Wing L., 1966). Автором этой работы было обнаружено, что внутрипарное сходство МЗ близнецов по частоте и амплитуде КГР выше, чем сходство ДЗ близнецов (показатели сходства оценивались с помощью внутрипарных корреляций). Высокое внутрипарное сходство МЗ по амплитудным и временным характеристикам КГР было обнаружено и в ряде других исследований, что свидетельствует о влиянии наследственных факторов на индивидуальные различия по этой характеристике (Rachman S., 1960; Lader M., Wing L., 1966; HumeW., 1973). КГР при орентировочной и оборонительной реакциях. Кроме показателей КГР в состоянии покоя (фоновых), в ряде работ изучалась КГР в экспериментальных ситуациях, вызывающих ориентировочную и оборонительную реакции.
Ориентировочная реакция (ОР) была впервые описана И.П.Павловым как двигательная реакция (поворот головы и глаз) на новый раздражитель, сопровождающаяся торможением текущей условно-рефлекторной деятельности. ОР имеет важное значение для организма, поскольку она, с одной стороны, выступает в качестве исследовательской реакции, а с другой — направлена на предупреждение возможных опасностей. Ориентировочная реакция на новый или неожиданный стимул представляет собой простейшую форму информационного поиска, тогда как оборонительная реакция (ОбР) — реакция организма на угрожающий стимул. Обе эти адаптивные реакции являются сложными функциональными системами и направлены на выживание и ориентировку организма в окружающей среде. И ОР, и ОбР сопровождаются множеством физиологических изменений, которые можно измерить, регистрируя электроэнцефалограмму, особенности сердечно-сосудистой деятельности и другие психофизиологические показатели. КГР представляет собой "ключевой" компонент ОР, отражающий значимость ситуации и "запускающий" ОР (Кочубей Б.И., 1983).
Имеются экспериментальные данные, свидетельствующие о том, что наследственные факторы лежат в основе индивидуальных различий показателей КГР при угашении ориентировочной реакции на звук и при оборонительной реакции, вызванной громким звуком. Для ОР и ОбР коэффициенты наследуемости составляют 1,0 и 0,62 соответственно (Zvolsky P. et al., 1976). Угашение ориентировочной реакции фиксируется при неоднократном повторении стимула, то есть при потере им качества новизны, и называется привыканием ОР.
В исследовании Б.И.Кочубея (1983, 1988) испытуемым предъявляли звуки различного тона и интенсивности, вызывающие либо ориентировочную, либо оборонительную реакции (По литературным данным, порог оборонительного компонента реакции лежит между 90 и 100 дБ, то есть можно считать, что звук интенсивностью 105 дБ, предъявлявшийся испытуемым, вызывал у них оборонительную реакцию.), и фиксировали различные параметры КГР (по Тарханову). Результаты, полученные Б.И.Кочубеем (1983, 1988), также подтвердили факт влияния генотипа на характеристики электрокожной активности при ОР и ОбР. Поскольку КГР является ключевым компонентом ОР, то есть отражает сам факт активации функциональной системы ОР, то, возможно, полученные данные позволяют говорить и о наследственной детерминации индивидуальных особенностей ОР.
Интересны также результаты изучения КГР в ситуации угашения ориентировочной реакции при повторении стимула — изучение "привыкания ОР". Согласно предложенной Е.Н.Соколовым (1960) концепции нервной модели стимула, ориентировочная реакция — это "информационный регулятор, включающийся при создании неопределенности и работающий в направлении получения информации, снимающей эту неопределенность" (Соколов Е.Н., 1964, с. 17). Нервная модель стимула, формирующаяся в процессе привыкания ОР, фиксирует огромное количество различных характеристик стимула. Каждый предъявляемый испытуемому стимул сличается с моделью. В случае рассогласования нервной модели стимула и параметров воспринимаемого стимула возникает ориентировочная реакция "что такое?", величина которой зависит от величины рассогласования. Привыкание ориентировочной реакции — проявление своеобразного механизма "информационного гомеостаза" (Гомеостаз — относительное динамическое постоянство внутренней среды организма; механизм поддержания гомеостаза обеспечивает адаптацию организма к изменяющимся условиям среды.).
Вообще привыкание, то есть способность высокоорганизованных организмов к прекращению ответных реакций на повторяющиеся незначимые стимулы, наряду со способностью реагировать на значимые или новые стимулы, является важнейшим механизмом адаптации, ведь в противном случае организму пришлось бы нести колоссальные энергетические затраты, и его выживание стало бы весьма проблематичным. Привыкание реакции на новый стимул можно рассматривать и как простейшую модель обучения. Неудивительно, что эта важнейшая характеристика связана с различными психологическими особенностями человека. Так, например, известно, что индивидуальные особенности ориентировочной реакции связаны с обучаемостью, когнитивными стилями, особенностями темперамента (Maltzman I., 1971;
Goldstein H.S. et al., 1970; Dronsejko K.,1972; Neary R.S., Zuckerman Z., 1976; Stelmack R.M., Plouffe L.,1983).
Поскольку явления ОР и ОбР имеют фундаментальный характер и являются одной из биологических систем адаптации, понятен и интерес генетиков поведения к этим реакциям, хотя их исследования немногочисленны.
Как показали работы M.Lader, L.Wing (1966), W.Hume (1973), Б.И.Кочубея (1983), D.Lykken с сотрудниками (1988), индивидуальные особенности привыкания КГР имеют выраженный наследственный компонент. По данным Б.И.Кочубея, вклад неаддитивных генетических факторов в межиндивидуальные различия по скорости привыкания КГР (Для оценки скорости привыкания, как правило, используют число попыток, предшествующих нулевым ответам (нет реакции), либо анализируют кривую привыкания, где по ординате дана величина электрокожной реакции, а по абциссе — номер попытки.) на звуки 80 (ОР) и 105 дБ (ОбР) составляет 74% и 53% соответственно.
Привыкание КГР исследовалось также на выборке разлученных близнецов (Lykken D. et al., 1994). Выборка испытуемых была достаточно большой: в исследовании приняли участие 43 пары разлученных МЗ и 25 пар разлученных ДЗ наряду с обычными близнецами (всего 79 пар МЗ и 42 пары ДЗ). Процедура исследования отличалась от традиционной экспериментальной парадигмы. При традиционном подходе к исследованию процесса привыкания испытуемому предъявляют ряд незначимых для него стимулов. Однако испытуемый может просто игнорировать стимулы, и за счет этого привыкание наступит быстрее; или, наоборот, испытуемый может специально обращать внимание на стимулы, например, считая их, что замедлит процесс привыкания. В результате вместо индивидуальных различий процесса привыкания экспериментатор может фиксировать, не подозревая об этом, различия в интерпретации задачи. Для решения этой проблемы В.Иконо и Д.Ликкен предложили давать испытуемым увлекательные задачи, требующие концентрации внимания, и при этом просили игнорировать незначимые, отвлекающие внимание стимулы. Этот подход хорошо зарекомендовал себя при дифференциации психически больных (Iacono W., 1982, 1983,1984) и здоровых людей. В.Иконо и Д.Ликкен (1983, 1984) показали, что, когда испытуемым предъявлялась определенная задача (например, слушать радиопередачу, смотреть фильм) и давалась инструкция игнорировать отвлекающие стимулы, привыкание происходило быстрее, по сравнению с ситуацией традиционно организованного эксперимента. Наблюдались значительные индивидуальные различия между людьми в электрической активности кожи: у одних испытуемых выраженная реакция на первые громкие звуки исчезала в конце серии звуков, у других реакция не исчезала и в конце серии, а у некоторых испытуемых реакции не было и на первые звуки.
В исследовании Д.Ликкена (1988) для регистрации максимальной реакции проводимости кожи (РПрК) использовалась следующая экспериментальная задача. Испытуемый должен был надувать воздушный шар, который находился внутри куба. Когда шар надувался до определенной величины, то иголка, расположенная в кубе, протыкала шар, и он лопался.
Максимум реакции определялся как пик реакции проводимости кожи во время выполнения этой задачи минус уровень проводимости кожи, зафиксированный перед выполнением экспериментальной задачи. Для оценки привыкания электрической активности кожи испытуемому во время прослушивания рассказа предъявляли серию громких звуков (105 дБ) с нерегулярным интервалом предъявления (от 20 до 100 сек).
Для того чтобы при генетическом анализе можно было сопоставлять данные разных испытуемых, Д.Ликкен использовал относительные оценки величин реакции КГР (в %) по отношению к диапазону вариации этой величины — от минимума реакции до максимума.
В психофизиологии, как и в большинстве областей психологического измерения, наблюдаемая переменная может являться коррелятом латентной психологической переменной. Часто предполагают, что функция, связывающая между собой наблюдаемую и латентную переменные, является монотонной и простой (например, линейной). Таким образом, если Y — психологическое действие стимула, то РПрК = а + b Y, где параметры а и b определяются физиологическими особенностями индивида. В частности, в случае привыкания КГР а будет равно нулю, поскольку мы полагаем, что при отсутствии ориентировочной реакции не должно быть реакции проводимости кожи, а коэффициент b будет отражать чувствительность периферических электрокожных мехнизмов к центральным изменениям, вызванным психологическим событием. Поскольку РПрК макс = bYмакс то при коррекции сырых данных (РПрК/РПрКмакс) элиминируется параметр b, который служит внешним (физиологическим) источником изменчивости. Действительно, "сырая" РПрК отражает как реактивность ЦНС на внешнее воздействие, так и эффекторную реактивность (зависящую, например, от числа потовых желез в области наложения электрода), тогда как скорректировать значения амплитуд отражают только активность ЦНС.
В результате коррекции сырых данных внутрипарные корреляции показателей КГР ДЗ близнецов увеличились до Уровня половины корреляции МЗ, что свидетельствует об аддитивном действии генов. Различия в данных, полученных для скорректированных и нескорректированных параметров, были обусловлены гетерогенностью последних.
Внутриклассовые корреляции показателей КГР оказались весьма высоки как у разлученных, так и у выросших вместе близнецов. Причем величина коэффициентов корреляции превышает ретестовую стабильность (Koriat A. et al., 1973; Iacono W. et al., 1984; Faulstich M. et al., 1986). Это свидетельствует о существенной роли наследственных факторов в индивидуальных различиях показателей КГР. Результаты метода подбора моделей также подтверждают вывод о большой роли генетических факторов (их действием можно объяснить от 21,7% до 70,3% фенотипической дисперсии указанного признака). Было выявлено значительное влияние фактора различающейся среды (в этот фактор входят также ошибка измерения и нестабильность признака), тогда как фактор общей среды (общая семья, школа, друзья и т.д.) оказался практически незначим для формирования индивидуальных особенностей показателей электрической активности кожи.
Итак, результаты немногочисленных исследований электрической активности кожи свидетельствуют о влиянии наследственных факторов на характеристики КГР как в состоянии покоя, так и при различных функциональных нагрузках, причем влияние генетических факторов при нагрузках больше.
|
|
|
|
|
Дата добавления: 2014-11-08; Просмотров: 1172; Нарушение авторских прав?; Мы поможем в написании вашей работы!