КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
История зоологии 2 страница
|
|
|
|
ПОДКЛАСС 1. КОРНЕНОЖКИ (Rhizopoda)
Корненожки характеризуются разнообразной формой, подвижными псевдоподиями и отсутствием дифференцировки цитоплазматического тела на более или менее постоянные зоны. В подклассе Rhizopoda различают 3 отряда: 1. Амебы (Amoebina). 2. Раковинные корненожки (Testacea). 3. Фораминиферы (Foraminifera).
Отряд 1. Амебы (Amoebina). Низшие, наиболее просто устроенные корненожки, лишенные скелета. Большинство амеб — обитатели пресных вод. Некоторые виды живут в море, а также в почве. Небольшое число — паразиты.
Строение и физиология. Размеры амеб различны: от 10—15 мкм до2—3 мм. Большинство амеб одноядерны, но есть и многоядерные виды. Рассмотрим в качестве примера обычную пресноводную амебу — протея (Amoeba proteus). Это довольно крупная амеба размером около 0,5 мм (рис. 1). Рассматривая ее под микроскопом, видим многочисленные разнообразной формы выросты — псевдоподии, или ложные ножки. Легко наблюдать, как форма псевдоподий все время меняется и благодаря этому происходит передвижение животного, которое обычно прикрепляется к субстрату. Происходит медленное перетекание амебы с места на место. Снаружи тело ее покрыто тонкой цитоплазматической мембраной (7,5 нм), за которой идет слой прозрачной довольно плотной эктоплазмы. Далее располагается зернистая полужидкая эндоплазма, составляющая основную массу тела животного.
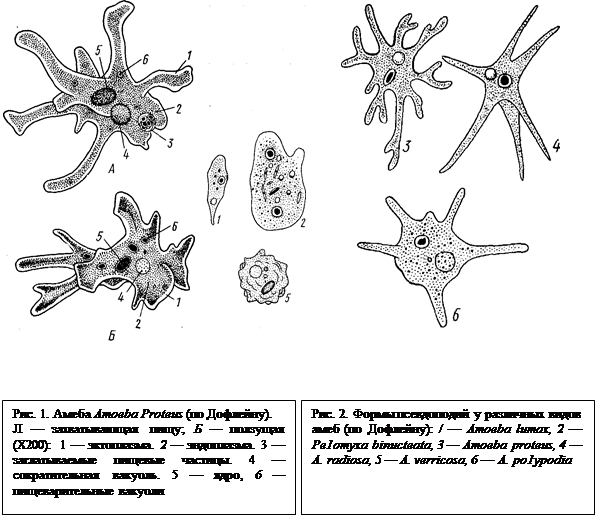 |
При формировании псевдоподии появляется сначала небольшой выступ эктоплазмы. Потом он растет и в него входит, как бы вливаясь, часть жидкой эндоплазмы. Псевдоподии амеб обычно не образуют разветвлений. Форма их может широко варьировать в зависимости от условий среды (рН, температура, содержание солей в воде и др.). Разные виды амеб отличаются по числу и форме псевдоподий (рис. 2). Последние могут быть короткими, длинными, тупыми, заостренными на концах и т. п. Быстрота движения Атоеbа proteus составляет при благоприятных условиях примерно 200 мкм в минуту.
|
|
|
Передвигаясь, амебы наталкиваются на различные мелкие объекты:
одноклеточные водоросли, клетки бактерий, частицы органического детрита, мелких простейших и т. п. Если объект достаточно мал, амеба обтекает его со всех сторон, и он вместе с небольшим количеством жидкости оказывается внутри цитоплазмы простейшего. Таким образом, в эндоплазме образуются пищеварительные вакуоли. Внутрь последних из эндоплазмы поступают пищеварительные ферменты, и происходит внутриклеточное пищеварение. Жидкие продукты переваривания поступают в эндоплазму. Амебы обладают ферментами, расщепляющими белки, углеводы и липиды. Вакуоль с непереваренными остатками пищи подходит к поверхности тела и выбрасывается наружу. У амеб нет ни постоянного рта, ни порошицы (отверстие для выведения экскрементов). У Атоеbа proteus пищей служат другие мелкие простейшие и одноклеточные водоросли. Описанный способ заглатывания оформленной пищи при посредстве псевдоподий называется фагоцитозом. Наряду с ним недавно (главным образом благодаря применению для исследования электронного микроскопа) открыт другой способ поступления веществ снаружи в тело амебы, который называется пиноцитозом. Этим путем поглощаются жидкие вещества. Снаружи внутрь цитоплазмы впячивается тонкий канал, имеющий форму трубочки, в которую засасывается окружающая амебу жидкость. Затем образующаяся пиноцитозная вакуоль отшнуровывается и оказывается лежащей внутри цитоплазмы. При помощи этого механизма амеба как бы «пьет» жидкость. Обычно пиноцитозные каналы и вакуоли очень малы (меньше микрометра) и поэтому их удается рассмотреть лишь на ультратонких срезах с применением электронного микроскопа. Кроме пищеварительных вакуолей в теле амеб находится еще одна так называемая сократительная, или пульсирующая, вакуоль. Это пузырек водянистой жидкости, который периодически нарастает, а затем, достигнув определенного объема, лопается, опорожняя свое содержимое наружу. Вскоре на том же месте снова появляется маленькая капелька, проделывающая тот же цикл. Промежуток между двумя пульсациями вакуоли у разных амеб равен 1—5 мин. Основная функция сократительной вакуоли — регуляция осмотического давления внутри тела простейшего. Вода из окружающей среды проникает в тело амебы через наружную мембрану осмотически. Концентрация различных растворенных веществ в теле амебы выше, чем в пресной воде, благодаря чему создается разность осмотического давления внутри и вне тела простейшего. Однако в организме простейшего есть своего рода откачивающий аппарат, периодически выводящий избыток воды из тела, — сократительная вакуоль. Подтверждением этой функции вакуоли служит распространение их преимущественно у пресноводных простейших. У морских и паразитических форм, окруженных жидкостью с более высоким, чем в пресной воде, осмотическим давлением, сократительные вакуоли обычно отсутствуют или же сокращаются очень редко.
|
|
|
Сократительная вакуоль кроме осморегуляторной функции частично выполняет и выделительную функцию, выводя вместе с водой в окружающую среду продукты обмена веществ. Однако основная функция выделения осуществляется непосредственно через наружную мембрану. Известную роль играет, вероятно, сократительная вакуоль и в процессе дыхания, ибо проникающая в результате осмоса в цитоплазму вода несет растворенный кислород.
В эндоплазме амеб находится пузырьковидное ядро, богатое ядерным соком и обладающее всеми компонентами клеточного ядра — оболочкой, пронизанной многочисленными порами, ядерным соком, хроматином и одним или несколькими ядрышками. Существуют некоторые виды амеб, обладающие не одним, а несколькими ядрами.
Многие виды свободноживущих амеб (особенно те, которые живут в почве) способны при наступлении неблагоприятных условий (например, подсыхании) инцистироваться. При этом амеба втягивает псевдоподии, округляется и выделяет вокруг себя прочную двойную оболочку белковой природы. В таком неактивном состоянии амебы могут сохраняться долгое время (месяцами), а затем вновь переходить к активной жизни.
|
|
|
Размножение. Амебам свойственно бесполое размножение, митотического деления ядра. Наблюдения авторов, считающих, что ядра амеб делятся путем амитоза, оказались ошибочными. На теле появляется легкий перехват, постепенно врезающийся в тело простейшего и перешнуровывающий ее на две дочерние особи (рис. 3). В период деления прием пищи обычно прекращается.
 |
Кишечные амебы человека и их значение. В кишечнике человека и ряда позвоночных обитает большое количество видов паразитических амеб, которые питаются содержимым кишечника, бактериями и большей частью не причиняют никакого вреда хозяину. Примером может служить кишечная амеба человека — Епtатоеbа coli (рис. 4). Однако среди обитающих в кишечнике человека амеб имеется один вид — дизентерийная амеба — Епtатоеbа histolytica, который может быть возбудителем тяжелой формы кишечного колита — амебиаза. Амеба эта имеет 20— 30 мкм в диаметре, подвижна (рис. 4,5).
 Живет она в толстых кишках человека и обычно питается бактериями, не нанося никакого вреда. Подобное явление, когда патогенный паразитический организм не проявляет своей патогенности, называется носительством. Но в ряде случаев дизентерийная амеба начинает вести себя иначе: проникает под слизистую оболочку кишки, начинает там питаться и усиленно размножаться. Слизистая кишечника изъязвляется, результатом чего бывает Распространение кишечных амеб осуществляется при помощи цист, выходящих вместе с фекальными массами наружу. Цисты очень стойки и длительное время сохраняют жизнеспособность и инвазионность (способность к заражению при попадании в кишечник человека). По строению цист можно установить вид амебы. Кишечная амеба Епtатоеbа coli (рис. 5) имеет восьмиядерные цисты, тогда как дизентерийная (Епtатоеbа histolytica) — четырехъядерные. В цистах есть особые ярко окрашивающиеся включения — хроматоидные тельца. При сильном заражении с экскрементами выводится до 300 млн. цист в день. Тяжелый кровавый понос (колит). Кишечные амебы человека распространены по всему Земному шару.
Живет она в толстых кишках человека и обычно питается бактериями, не нанося никакого вреда. Подобное явление, когда патогенный паразитический организм не проявляет своей патогенности, называется носительством. Но в ряде случаев дизентерийная амеба начинает вести себя иначе: проникает под слизистую оболочку кишки, начинает там питаться и усиленно размножаться. Слизистая кишечника изъязвляется, результатом чего бывает Распространение кишечных амеб осуществляется при помощи цист, выходящих вместе с фекальными массами наружу. Цисты очень стойки и длительное время сохраняют жизнеспособность и инвазионность (способность к заражению при попадании в кишечник человека). По строению цист можно установить вид амебы. Кишечная амеба Епtатоеbа coli (рис. 5) имеет восьмиядерные цисты, тогда как дизентерийная (Епtатоеbа histolytica) — четырехъядерные. В цистах есть особые ярко окрашивающиеся включения — хроматоидные тельца. При сильном заражении с экскрементами выводится до 300 млн. цист в день. Тяжелый кровавый понос (колит). Кишечные амебы человека распространены по всему Земному шару.
|
|
|
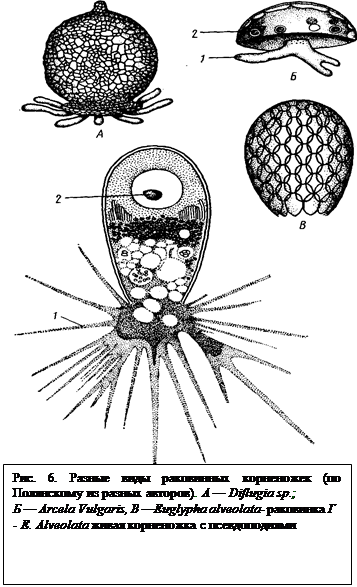
 Отряд 2. Раковинные амебы (Теstасеа). Представители этого отряда отличаются от амеб защитной раковиной, одевающей тело. Раковина Теstасеа обычно имеет вид округлого или овального мешочка с отверстием (устьем), из которого выдаются псевдоподии, имеющие у разных видов неодинаковую форму и длину. Раковина у одних форм представляет тонкий слой плотного органического вещества, у других она образуется посторонними частицами (песчинки и т. п.), склонными выделениями цитоплазмы (рис. 6).
Отряд 2. Раковинные амебы (Теstасеа). Представители этого отряда отличаются от амеб защитной раковиной, одевающей тело. Раковина Теstасеа обычно имеет вид округлого или овального мешочка с отверстием (устьем), из которого выдаются псевдоподии, имеющие у разных видов неодинаковую форму и длину. Раковина у одних форм представляет тонкий слой плотного органического вещества, у других она образуется посторонними частицами (песчинки и т. п.), склонными выделениями цитоплазмы (рис. 6).
Размножение раковинных амеб совершается, как и у представителей предыдущего отряда, делением надвое, причем одна из половин выходит наружу через устье и окружается новой раковиной (рис. 7).
Раковинные корненожки распространены в пресных водах, встречаются преимущественно среди прибрежной растительности, на дне вблизи берега. Значительное число видов Testacea живет в торфяных болотах.
Отряд 3. Фораминиферы (Foraminifera). Строение и физиология. Фораминиферы, обитатели моря, устроены сложнее прочих корненожек. Число современных видов превышает 1000. Раковина фораминифер обнаруживает ряд этапов постепенного усложнения.
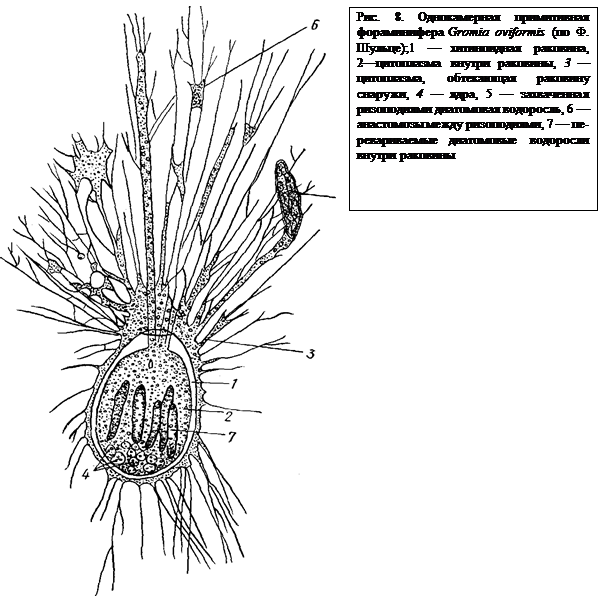
В наиболее простых случаях раковина состоит из плотного органического вещества — псевдохитина (рис. 8). Это вещество выделяется эктоплазмой. У других видов к этой тонкой пленке приклеиваются захваченные псевдоподиями посторонние частицы, главным образом песчинки. Получается хитиноидная основа, инкрустированная зернами кварца. Раковинки подобного типа массивны и тяжеловесны (например, роды Hyperammina, Rhabdammina, Astorhize, рис. 9,3—7). У большинства современных фораминифер раковина тоже тонкой хитиноидной основы, но пропитанная углекислым кальцием. Обладая большой прочностью, раковины такого рода отличаются гораздо большей легкостью, чем инкрустированные.
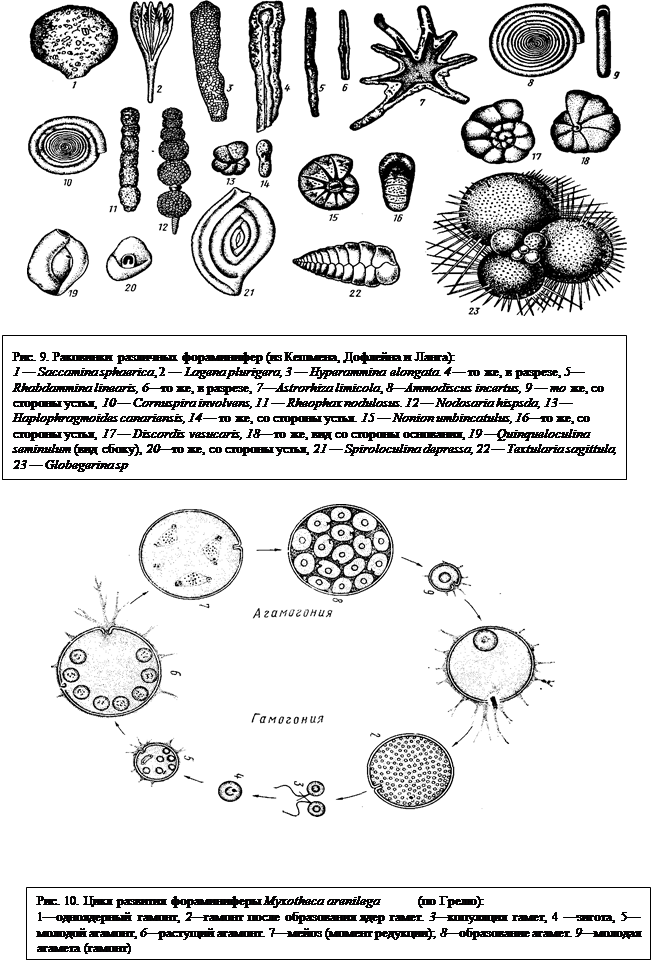
Форма раковины фораминифер чрезвычайно разнообразна (рис. 9). У некоторых видов раковина имеет форму продолговатого мешка, у других вытягивается в трубку, у третьих эта трубка закручивается в спираль. Все это однокамерные формы (рис. 9, 1, 2, 8, 9, 10). Но у большинства фораминифер полость раковины поделена поперечными перегородками на камеры (многокамерные формы), которые сообщаются друг с другом отверстиями, имеющимися в перегородках. Взаимное расположение камер может быть различным (рис. 9). Они могут располагаться в один ряд, в два ряда, спирально и т. п. Каждая многокамерная корненожка начинает свою жизнь будучи однокамерной, причем эта первая камера меньше позднейших и называется зародышевой. Отверстие, сообщающее раковину с внешним миром и служащее для выхода псевдоподий, называется устьем. Помимо устья у многих корненожек все стенки раковины пронизаны тончайшими порами, тоже служащими для выхода ложноножек. Строение псевдоподий у Foraminifera, которые называются ризоподиями, чрезвычайно своеобразно. Они представляют собой длинные тонкие переплетающиеся и сливающиеся нити (см. рис. 8), образующие вокруг раковинки сложную сеть. В ризоподиях осуществляется непрерывный ток цитоплазмы. По одной и той же ризоподии одни струи ее текут в центростремительном (к раковине), другие — в центробежном направлениях. Ризоподии служат для улавливания и, частично, переваривания пищи, а также для передвижения животного. Они способны сокращаться и вытягиваться
Размножение. Чередование поколений. Фораминиферы обладают сложным жизненным циклом, в который включаются две формы размножения — бесполое и половое. Некоторые стороны жизненного цикла их были изучены лишь за последнее время. Рассмотрим в качестве примера цикл развития однокамерной корненожки Myxotheca arenilega (рис. 10).
Описание цикла начнем с одноядерной стадии (рис. 10, 1), которая называется гамонтом, Это название связано с ее дальнейшей судьбой. После некоторого периода свободной активной жизни ядро ее начинает многократно делиться, и корненожка становится многоядерной (рис. 10. 2). Вокруг каждого из ядер обособляется небольшой участок цитоплазмы, и все тело корненожки распадается на множество мелких клеток, каждая из которых формирует два жгутика неравной длины. Клетки выходят из раковины наружу в морскую воду и попарно сливаются (рис. 10, 3). Таким образом, это половые клетки (гаметы), в результате слияния которых образуется зигота (рис. 10, 4). Она дает начало новому поколению, формирующему раковину и называемую агамонтом (рис. 10, 5, 6). В агамонте происходит постоянное увеличение числа ядер в результате их митозов, и сам он увеличивается в размерах. Зигота обладает диплоидным комплексом хромосом, так же как и развивающийся из зиготы агамонт. После завершения его роста происходит еще два деления ядер (рис. 10, 7), которые являются редукционными (мейоз), и образующиеся в результате их ядра гаплоидны. Далее вокруг каждого ядра вновь обособляется участок цитоплазмы и все тело распадается на большое количество мелких одноядерных зародышей, именуемых агаметами (частица «а» по-гречески означает отрицание, так что их можно было бы назвать «негаметами»). Это бесполое размножение, которое ведет к образованию нового поколения с гаплоидным ядром. Каждая агамета окружается раковиной и дает начало гамонту — поколению, с которого мы начали рассмотрение цикла и который далее вновь образует гаметы. Таким образом, в жизненном цикле фораминиферы чередуются две формы размножения: половое (при посредстве гамет) и бесполое (через агаметы) и два поколения: гамонты (размножаются половым путем) и агамонты (размножаются бесполым путем).
Особенно замечательно в жизненном цикле фораминифер относительно недавно открытое чередование гаплоидной и диплоидной фаз. Мы видели, что редукция (мейоз) происходит здесь не перед образованием гамет, как это бывает у всех многоклеточных животных, а при образовании стадий бесполого размножения—агамет. Это единственный случай в животном мире, когда одно поколение гаплоидно, а другое диплоидно. Напротив, в растительном мире, где закономерно чередуются спорофит и гаметофит, происходит также и чередование гаплоидной (гаметофит) и диплоидной (спорофит) фаз ядра. Разумеется, это сходство не говорит о связи фораминифер с растениями, но представляет интересный пример конвергентного развития.
В деталях (строение гамет, их число, судьба гамонтов и т. п.) жизненные циклы фораминифер очень разнообразны, и мы не можем останавливаться на их подробном рассмотрении. Однако у всех изученных видов он слагается из двух поколений — полового и бесполого.
Большинство Foraminifera живет на дне водоемов, иногда на глубинах в тысячи метров, питаясь разными мелкими организмами. Лишь немногие виды, например Globegerina, входят в состав планктона. Раковинки этих видов снабжены обычно длинными радиальными шипами, сильно увеличивающими поверхность и позволяющими «парить» в толще воды.
Палеонтология и практическое значение. Корненожки входят в состав очень многих морских отложений, начиная с древнейших кембрийских. Толстые слои известняков, мела, зеленого песчаника и некоторых других осадочных пород состоят преимущественно из раковин фораминифер.
Благодаря малым размерам и большой распространенности корненожки во многих отношениях удобнее, чем остатки крупных организмов, для определения возраста геологических пластов при поисках нефти и других полезных ископаемых. Каждый пласт имеет свою характерную фауну корненожек. Поэтому в каждом нефтеносном районе достаточно один раз установить залегание нефти в определенном пласте для того, чтобы и в других местах узнавать о вероятной близости нефти по исследованию корненожек в пробах пород, добываемых из буровых скважин. В СССР, США и других странах имеются специальные лаборатории для изучения распределения корненожек в нефтеносных слоях. Этот метод широко используется в практике геологоразведочных работ.
. ПОДКЛАСС II. ЛУЧЕВИКИ (RADIOLARIA)
Строение и физиология. Сюда относятся Sarcodina, отличающиеся геометрической правильностью и необычайным разнообразием форм, что связано с наличием у них внутреннего скелета. Лучевики — исключительно морские животные, ведущие планктонный образ жизни. К этому подклассу относится 7—8 тыс. видов. Они встречаются на всех глубинах. Советские экспедиции находили радиолярий на глубинах, превышавших 8000 м. Наиболее многочисленны они в теплых морях.
Большинство радиолярий обладает минеральным сложно устроенным внутренним скелетом. Тело их в большинстве случаев шарообразно, но может быть и другой формы. От тела во все стороны отходят многочисленные нитевидные, частично анастомозирующие псевдоподии (рис. 11). Большинство лучевиков обладает одним крупным центрально расположенным ядром, но встречаются и многоядерные формы.
 |
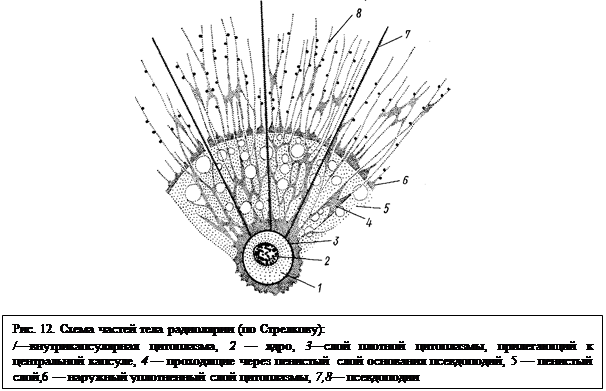
Ядро, занимающее, как правило, срединную часть клетки, окружено зоной гомогенной и довольно плотной внутрикапсулярной цитоплазмы
(рис. 12). Дело в том, что эта часть тела радиолярии заключена в прочную центральную капсулу; последняя представляет собой пронизанную отверстиями (чаще многочисленными), состоящую из органического вещества кожистую мембрану, похожую, таким образом, на органическую раковину. Лишь у одного отряда Acanaria центральная капсула состоит из слоя уплотненной цитоплазмы или может даже совсем отсутствовать. Через отверстия внутрикапсулярная цитоплазма сообщается с ее более наружными частями — внекапсулярой цитоплазмой. Это разделение цитоплазмы радиолярий на две зоны не соответствует обычному для простейших делению на эктоплазму и эндоплазму и возникло в пределах подкласса лучевиков независимо от других групп простейших. Во внекапсулярой цитоплазме возникает дальнейшая дифференцировка (рис. 12).
Непосредственно к центральной капсуле прилежит слой зернистой плотной цитоплазмы. За ней кнаружи следует обычно очень мощный слой пенистой цитоплазмы с многочисленными включениями — слизистыми, жировыми и т. п. Кнаружи от этого слоя расположен тонкий периферический слой сетчатой цитоплазмы, непосредственно граничащий с наружной средой. Радиолярии обычно имеют псевдоподии двоякого характера. Одни из них берут начало от самого глубокого слоя внекапсулярной цитоплазмы, проходят через пенистый слой и выдаются наружу. Другие же непосредственно связаны с самым наружным слоем цитоплазмы. Именно эти псевдоподии образуют, анастомозируя друг с другом, сложную сеть, играющую основную роль в улавливании пищи (водоросли, другие простейшие и т. п.). Кроме описанных форм псевдоподий у многих лучевиков есть еще прямые не ветвящиеся аксоподии (напоминающие аналогичные образования у солнечников, с. 35), обладающие осевой скелетной нитью и расположенные более или менее строго радиально. Эти псевдоподии увеличивают общую поверхность тела простейшего, что способствует «парению» в воде. К аксоподиям также могут прилипать пищевые частицы.
В цитоплазме лучевиков часто имеются одноклеточные симбиотические водоросли — зеленые (зоохлореллы) или желтоватые (зооксантеллы). Эти симбионты снабжают лучевика кислородом, сами же частично перевариваются и служат источником питания.
Лишь немногие лучевики лишены скелета. У большинства имеется сложно устроенный минеральный скелет, состоящий из окиси кремния или же из сернокислого стронция (SrSO4) (отр. Acantharia). В скелете радиолярий сочетаются прочность и легкость и выполняет он двоякую функцию — защищает тело простейшего и способствует парению в воде в результате увеличения поверхности путем образования различных выростов, например многочисленных игл. Часто скелет геометрически
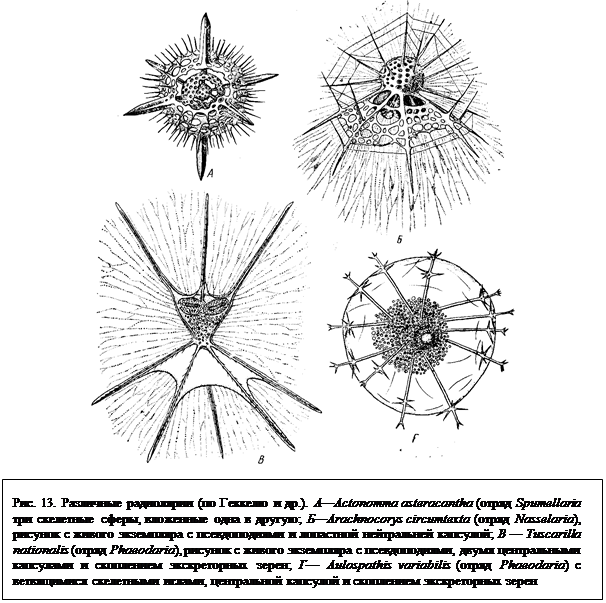
правильной формы (рис. 13). В нем могут сочетаться отдельные правильно расположенные иглы (спикулы), полые ажурные шары, вложенные друг в друга, различные, подчас причудливой формы образования, напоминающие колокольчики, головные уборы, короны и т. п. (рис. 13).
Размножение. До настоящего времени формы размножения радиолярий изучены еще недостаточно. Многие виды размножаются бесполым путем, делясь надвое. При этом скелетные элементы либо распределяются между дочерними особями, либо у одной из дочерних особей скелет формируется заново. Ядро, расположенное в центральной капсуле, при этом делится надвое. Во время этого деления ядра наблюдается необычно большое число хромосом, превышающее 1000. В последнее время доказано, что большое число хромосом зависит от высокого уровня полиплоидности ядра. Подобные высокополиплоидные ядра радиолярий называются первичными. У немногих видов радиолярий, например у Aulacantha scolymantha из отряда Phaeodaria, наблюдался процесс распада первичного ядра на большое число вторичных ядер, обладавших небольшим числом хромосом. Вокруг каждого вторичного ядра обособляется участок цитоплазмы и получается стадия бесполого размножения, соответствующая агамете фораминифер Из такой агаметы далее развивается взрослая радиолярия, а в ядре происходит умножение числа хромосом и превращение его в первичное высокополиплоидное ядро. Процесс этот, однако, полностью еще не прослежен. Что касается полового процесса, то он известен пока лишь у немногих представителей отряда Acantharia. При этом тело радиолярий распадается на множество мелких двужгутиковых гамет, сливающихся затем попарно. Из зиготы развивается взрослая акантария.
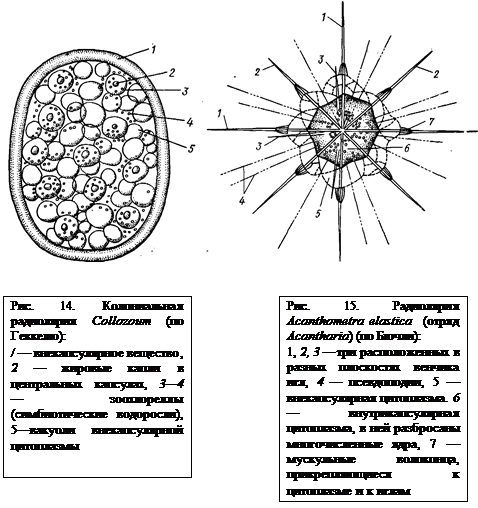
Колонии. Громадное большинство Radiolaria одиночны, но у некоторых форм образуются шаровидные или колбасовидные колонии (рис. 14), содержащие в общей массе внекапсулярного вещества не одну, а множество центральных капсул, каждая из которых отвечает отдельной особи. Такие колонии образуются из единственной первичной особи путем деления ядер и центральных капсул, тогда как внекапсулярная цитоплазма только растет, не принимая участия в делении. Колонии среди отряда Spumellaria могут достигать значительной величины— 1,5—2,0 см.
 |
Классификация. В пределах подкласса радиолярий различают пять отрядов.
Отряд 1. Acantharia. Основу скелета акантарий, слагающегося из SrSo4, составляют 20 радиально расположенных игл (рис. 15), сходящихся в центре животного. Иглы образуют 5 поясов по 4 иглы в каждом. Свободные концы игл торчат из тела простейшего наружу. Разная степень развития игл создает большое многообразие форм в пределах отряда. Скелетные иглы прикрепляются к наружному слою цитоплазмы при помощи мускульных волоконец, способных к сокращению. При их сокращении или удлинении меняется общий объем тела простейшего. Это механизм очень тонкой «настройки» для поддержания акантарий в состоянии парения в толще морской воды. Acantharia обладают центральной капсулой, состоящей из уплотненной цитоплазмы. Наиболее примитивные представители этого отряда лишены центральной капсулы. Представитель — Acanthometra elastica (рис. 15).
Отряд 2. Spumellaria. Единичные виды отряда лишены скелета (см. рис. 11), но у большинства имеется разнообразно устроенный кремнеземный скелет. Наиболее примитивной и, вероятно, исходной формой служат мелкие иглы, разбросанные тангенциально в эктоплазме. В результате слияния игл образуются шары, нередко вложенные друг в друга и соединенные радиально идущими иглами. Центральная капсула с многочисленными порами. Представители: Thalassicola muleata, Chromyodrymus abiatanus.
Отряд 3. Nasselaria. Представители этого многочисленного отряда характеризуются очень разнообразным кремнеземным скелетом. Основой здесь является четырехлучевая спикула, к которой могут присоединяться дополнительные кольца. На этой основе возникают разнообразные подчас причудливые формы скелетов в виде ажурных шапочек, шлемов, шаров и т. п. (см. рис. 13). Центральная капсула нередко не сферическая, а вытянутая в длину, веретеновидная. Представитель — Medusetta craspedota.
Отряд 4. Phaeodaria. В высшей степени разнообразно устроен кремневый скелет представителей этого отряда, к которому относятся наиболее глубоководные виды (рис. 13). У некоторых имеются створки, окружающие центральную капсулу. Встречаются также разнообразные радиальные, нередко ветвящиеся иглы. Характерным признаком отряда служит строение центральной капсулы с одним или тремя крупными отверстиями вместо многочисленных пор. Характерно также наличие во внекапсулярной цитоплазме в области центрального отверстия капсулы особого образования, именуемого феодиумом. Последний обычно окрашен пигментом в темный цвет и представляет скопление выделительных телец, а также пищевых включений и резервных веществ. Он всегда резко выделяется на фоне бесцветной цитоплазмы. Представитель — Aulacantha scolymantha.
Отряд 5. Sticholonchea. Своеобразные радиолярии, представленные пока единственным родом Sticholonche, найденным в Тихом океане и Средиземном море. Характеризуется вытянутым двусторонне-симметричным телом и наличием радиальных игл, расположенных 18—20 пучками.
ПОДКЛАСС III. СОЛНЕЧНИКИ (HELIOZOA)
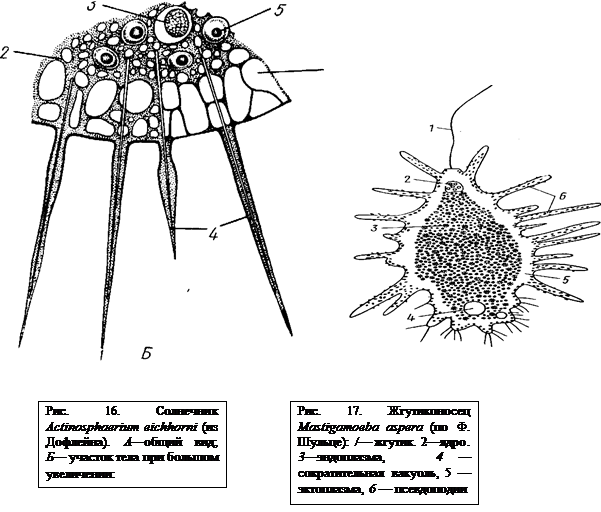
К солнечникам относятся всего несколько десятков видов пресноводных и морских простейших. От радиолярий они отличаются отсутствием центральной капсулы. Тело, чаще всего шаровидное, распадается на широкую зону эктоплазмы и эндоплазму (рис. 16). Имеются многочисленные радиально расположенные аксоподии (с. 32). В эндоплазме одно или несколько ядер. В эктоплазме пресноводных видов есть редко сокращающиеся пульсирующие вакуоли. Большинство видов лишено минерального скелета. Некоторые морские солнечники имеют решетчатый сферический скелет из кремнезема. Бесполое размножение путем деления. У некоторых видов описан половой процесс. Представитель Actinosphaerium eichhorni.
КЛАСС II. ЖГУТИКОНОСЦЫ (MASTIGOPHORA)
Класс жгутиконосцев характеризуется наличием жгутов, служащих органоидами движения. Их может быть один, два или множество. Мы видели, что и у саркодовых на известных стадиях жизненного цикла (у гамет) имеются жгутики. Но у них жгутиконосные стадии преходящи, тогда как у Mastigophora жгутики присутствуют постоянно в течение большей части жизненного цикла. Наряду с этим у немногих видов жгутиконосцев могут временно или постоянно присутствовать и псевдоподии (рис. 17). Все это указывает на близость, и даже отсутствие резкой границы между саркодовыми и жгутиконосцами, что дает основание объединять эти два класса в тип Sarcomastigophora.
 Жгутиконосцы представляют большой интерес в том отношении, что в пределах этого класса проходит как бы граница между растительным и животным миром. Представители ряда групп жгутиконосцев обладают хроматофорами, содержащими хлорофилл. Эти формы, как настоящие зеленые растения, способны на свету осуществлять фотосинтез. Другим жгутиковым свойствен гетеротрофный обмен — как все животные, они используют в качестве пищи готовые органические вещества. Имеются, наконец, виды, совмещающие обе формы обмена. Поэтому мы рассмотрим весь класс жгутиконосцев в целом, не выделяя из него растительные формы.
Жгутиконосцы представляют большой интерес в том отношении, что в пределах этого класса проходит как бы граница между растительным и животным миром. Представители ряда групп жгутиконосцев обладают хроматофорами, содержащими хлорофилл. Эти формы, как настоящие зеленые растения, способны на свету осуществлять фотосинтез. Другим жгутиковым свойствен гетеротрофный обмен — как все животные, они используют в качестве пищи готовые органические вещества. Имеются, наконец, виды, совмещающие обе формы обмена. Поэтому мы рассмотрим весь класс жгутиконосцев в целом, не выделяя из него растительные формы.
Класс Mastigophora распадается на два подкласса: 1. Phytomastigina (растительные жгутиконосцы, способные к фотосинтезу) и 2. Zoomastigina (животные жгутиконосцы с гетеротрофным типом обмена).
Строение и физиология. Размеры и форма тела жгутиконосцев довольно разнообразны: оно часто бывает яйцевидным, цилиндрическим, шаровидным, бутылковидным и т. п. Иногда, как у представителей отр. Dinoflagellata, тело может приобретать различные выросты и принимать причудливую форму (рис. 18). Цитоплазма делится на экто- и эндоплазму. У некоторых Mastigophora цитоплазма снаружи ограничена лишь элементарной мембраной (у тех немногих видов, которые способны образовывать псевдоподии). У других наружный слой эктоплазмы уплотняется и образует пелликулу, в результате чего тело простейшего теряет способность к изменению формы. Наконец, у многих жгутиконосцев на поверхности клетки выделяется специальная оболочка; последняя у форм, относящихся к разным

отрядам, может состоять из различных веществ: из хитиноидного органического вещества, из студенистого вещества (у многих колониальных видов) или же, как у типичных растительных клеток, из клетчатки (у растительных жгутиконосцев).
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 2730; Нарушение авторских прав?; Мы поможем в написании вашей работы!