КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Возникновение приспособлений (адаптациогенез) 2 страница
|
|
|
|
Многие морфологические приспособления были бы совершенно непонятны, если бы мы не знали их экологического и, в частности, биоценотического значения. Таковы взаимные приспособления организмов. Как раз в этих случаях нелегко себе представить адаптивность ничтожных различий во взаимных приспособлениях различных форм одного вида или совершенно различных организмов. Трудность увеличивается здесь именно необходимостью взаимного согласования этих изменений. В особенности это касается половых различий. И все же половые части многих насекомых в высшей степени пластичны и всегда хорошо согласовываются в обоих полах. Приходится и здесь допустить, что естественный отбор имеет дело только с очень малыми изменениями, которые не вносят существенных нарушений в половые взаимоотношения.
Не столь очевидно строги требования взаимного приспособления цветов и насекомых-опылителей, поскольку узкая специализация наблюдается здесь довольно редко. Нетрудно понять, что даже небольшое удлинение хоботка пчелы может иметь приспособительное значение, позволяя использовать для добычи меда также растения с более глубокими нектарниками К Четыре вида небольших жуков рода Diharda незначительно отличаются друг от друга по своим размерам и окраске. Трудно было бы признать адаптивное значение этих ничтожных различий, если бы не было известно, что они относятся к мирмекофилам и, живя в муравейниках, соответствуют по размерам и окраске четырем различным видам муравьев, среди которых они живут.
Если существуют известные трудности для изучения возникновения новых приспособлений в природной обстановке, то это восполняется рядом очень интересных наблюдений над нашими культурами. Возникновение морфологических различий экологического характера можно прекрасно проследить на истории животных и растений в искусственных условиях и, в особенности, на истории вредителей сельского хозяйства и сорняков полевых культур, которые показывают приспособление организмов к новым условиям, создаваемым человеком.
|
|
|
Уже исследования Веттщтейна над влиянием покоса на растительность альпийских лугов показали наличие значительных изменений характера «приспособления» к деятельности человека. Подобные исследования были затем предприняты Н. В. Цингером, который показал, что, например, горный вид погремка (Alectorolophus montanus), цветущий на нетронутых альпийских лугах в течение всего лета, дал на укосных лугах начало рано зацветающему летнему виду (A. aestivalis). В условиях двух покосов этот вид разбился на два — раннеспелый весенний вид (A. vernalis) и позднеспелый летний (A. polycladus). Еще интереснее, что в условиях посевов ржи летняя форма преобразовалась в особую форму, приспособившуюся к этим посевам (A. apterus). У этой формы коробочки не раскрываются и семена сами не осыпаются (они требуют обмолота!). Крылья, бывшие на семенах исходных форм, редуцировались, и поэтому семена не отвеиваются. Они остаются в урожае ржи. Таким образом этот вид погремка превратился в сорняк. Эволюция сорняка, однако, еще не вполне закончена. У этой формы (A. apterus) еще сохранились признаки, указывающие на ее происхождение: иногда попадаются коробочки с щелью, через которую семена еще могут высыпаться; среди семян часто встречаются, кроме бескрылых, также и семена с узкой оторочкой—рудиментом крыльев (52% семян) и в небольшом числе (12%)—с заметными или даже (1,5%) хорошо развитыми крыльями.
В условиях систематического покоса происходит также отбор низкорослых особей растений — так, например, устанавливается низкорослая форма лебеды, не уничтожаемая при кошении. Интересны и другие исследования Н. Цингера над происхождением сорняков и в особенности над растениями, обычно засоряющими посевы льна. Льняной рыжик (Camelina linicola) близок и, очевидно, произошел от ярового рыжика (С. glabrata), отличаясь от него прежде всего более крупными семенами, которые по весу приближаются к семенам льна. То же самое касается льняной гречихи {Polygonum linicola), происходящей от дикой P. lapathifoliurn, льняной торицы (Spergula linicola), происходящей от обыкновенной S. vulgaris, льняного плевела (Lolium remotum), происходящего от обыкновенного сорняка яровых посевов L. temulentum. Происхождение этих форм связано с бессознательным отбором, так как при просеивании семян льна всегда оставались самые тяжелые семена этих растений, наиболее приближающиеся по весу к семенам льна. Вес семян был ведущим морфологическим признаком, по которому шло «приспособление» новых видов-сорняков к условиям жизни и размножения в посевах льна.
|
|
|
Новейшие данные, касающиеся происхождения культурных злаков, показывают, что рожь и овес вошли, очевидно, в человеческую культуру первоначально как сорняки посевов пшеницы и ячменя. По мере продвижения культуры на север они все более засоряли посевы, но вместе с тем приобретали все большее значение, сначала как непроизвольная и безвредная примесь, а затем как самостоятельный объект культуры.
Наконец, и вообще история культивируемых растений и домашних животных дает картину очень постепенного физиологического и морфологического «приспособления» этих организмов к своеобразным экологическим условиям, создаваемым человеком. Далеко не все эти изменения направляются человеком. Последнего интересуют всегда лишь очень немногие свойства. Остальные сопутствующие изменения могут иметь характер обычного «приспособления», либо иногда (чаще, чем в природной обстановке) характер действительно вполне «индифферентных» признаков.
3. ВОЗНИКНОВЕНИЕ АДАПТИВНЫХ ФОРМООБРАЗОВАТЕЛЬНЫХ РЕАКЦИЙ
Хотя и рассмотренные физиологические и морфологические приспособления развиваются на основе измененной нормы реакций организма, вопрос о возникновении способности к адаптивным реакциям организма все же гораздо сложнее, чем рассмотренные вопросы, так как здесь речь идет не об одной типичной мор-фогенетической реакции, а о возможности целого ряда таких реакций, сохраняющих свою адаптивность при изменении факторов внешней среды и изменении положения организма в этой среде.
|
|
|
Вместе с тем этот вопрос имеет большое общее значение, так как индивидуальная приспособляемость организма создает для него особое положение весьма гибкой устойчивости, т. е. определенной «лабильности», позволяющей ему не только переживать периоды значительных и резких изменений в факторах среды, но и активно переходить из одной среды в другую и даже перестраивать свою организацию. Поэтому проблема развития системы адаптивных реакций организма имеет большое значение и для понимания закономерностей эволюционного процесса.
Кроме того, она относится к числу крайне мало разработанных вопросов эволюционной теории, так как ламаркисты исходили в своих построениях из фактов индивидуальной приспособляемости как заранее данной предпосылки, не вникая в вопрос об ее возникновении. Неодарвинисты же не придавали ему значения, так как предполагалось, что результаты индивидуального приспособления, не будучи наследственными, не имеют значения в процессе эволюции. Мы здесь рассматриваем эти вопросы только в отношении морфогенетических реакций.
А. ЭЛЕМЕНТАРНЫЕ ЗАВИСИМОСТИ РЕАКЦИИ И ИХ ПРЕОБРАЗОВАНИЕ
а. Зависимое развитие морфозов. Элементарные формы реагирования мы усматриваем в тех изменениях индивидуального формообразования, которые наступают в связи с изменениями в факторах внешней среды и не имеют еще под собой заметной исторической базы, а могут рассматриваться как новые реакции. Такие изменения мы называем морфозами, Если они оказываются сходными с известными мутациями, то их называют также фенокопия-ми. Морфозы вызываются чаще всего действием необычных факторов — лучей Рентгена, химических соединений или обычными факторами при необычайной их интенсивности, например, крайне высокими или крайне низкими температурами. В таких морфозах нет ничего приспособительного — это такие же нарушения развития, как и видимые мутации. Большого интереса для нас они здесь не имеют. В разбираемом вопросе генезиса адаптивных модификаций имеют значение главным образом лишь такие реакции или морфозы, которые связаны с обычными в нормальной среде изменениями внешних факторов. Такие морфозы отличаются не столь резким выражением, они не имеют такого «патологического» характера, как обычные рентгено-хемо- или термоморфозы. Однако частично именно поэтому они представляют особый интерес.
|
|
|
Известна зависимость выражения многих мутаций от изменений внешних факторов. Некоторые мутации, как, например, мутация безглазия (eyeless) у дрозофилы или мутация уродливого брюшка (Abnormal abdomen) получают свое полное выражение лишь при развитии личинок в свежей питательной среде; на подсохшем корму развиваются мухи, приближающиеся по своему фенотипу к дикой норме. Другие мутации зависят в своем выражении от температурных условий. Мутация удвоения конечностей (reduplicated) дрозофилы получает свое типичное выражение лишь при низкой температуре. При обычной температуре культивирования личинок ее выражение исчезает. Интересно, что для близких мутаций такие зависимости могут иметь прямо противоположный характер. Так, мутация рудиментарных крыльев (vestigial) дрозофилы получает свое наиболее ясное выражение при низких температур культивирования; по мере повышения этой температуры размеры крыльев увеличиваются, и при 32° у известного числа мух развиваются почти нормальные крылья (рис. 22). С другой стороны, аллеломорфная (по отношению к vestigial) мутация pennant, обладающая также редуцированными крыльями, получает свое полное выражение только при высокой -температуре культивирования. По мере понижения этой температуры в пределах от 30 до 16о размеры крыльев увеличиваются и достигают нормы (рис. 23). У двух аллеломорфных мутаций температурная реакция тканей одного и того же зачатка органа изменяется в противоположных направлениях. Нормальный же аллеломорф этих мутаций отличается максимальной устойчивостью и дает лишь слабую температурную реакцию. Интересно, что оба мутантных аллеломорфа вместе (компаунд vestigial X pennant) дают закономерную смену направления реакции: при повышении температуры в пределах от 16 до 22° культивирования величина крыльев уменьшается (как у pennant), при повышении температуры от 22 до 26° размеры крыльев остаются неизменными { морфогенез устойчив), а при дальнейшем поднятии температуры от 26 до 32е крылья увеличиваются (как у vestigial; см. рис. 24, табл. 10. Это показывает, что характер реакции зависит не от отдельного «гена», а от их комбинации и от всего генотипа в целом. Мы уже обращали внимание на то, что наиболее изменчивым выражением обладают многие рецессивные мутации в гомозиготе (например, eyeless, vestigial, pennant, reduplicated) и полудоминантные мутации в гетерозиготе (напр., Bar, Abnormal abdomen). Это касается как их чувствительности к изменениям в факторах внешней среды, так и легкой изменяемости реакций и форм реагирования в зависимости от незначительных изменений генотипа (при комбинировании малых мутаций — модификаторов). Поэтому и форма реагирования может быть изменена не только путем внесения второй мутации (как в примере vestigial X pennant), но и в результате искусственного (а следовательно — и естественного) отбора малых изменений, зависящих от небольших различий в генотипе (рис. 25).
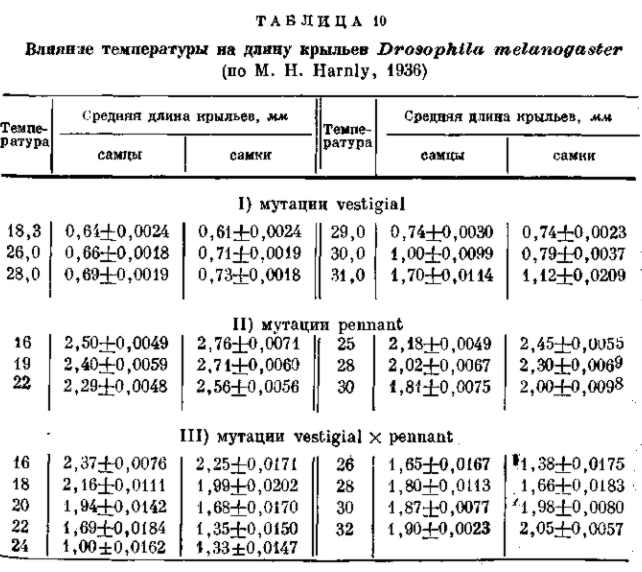
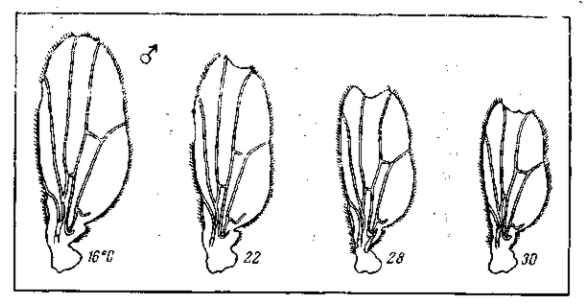
Рис. 23. Зависимость формы и величины крыльев самцов мутации pennant (Drosophila melanogaster) от температуры. Эта мутация показывает реакцию, обратную по сравнению с ее аллеломорфом vestigial. В этом случае при низкой температуре 16° С около 13% особей имеют совершенно нормальные крылья. По М. Г. и М. Л. Харнли, 1935
Экспериментов, показывающих эффективность искусственного отбора на усиление или ослабление выражения известных мутаций, было проведено очень много. Нас интересует здесь, однако, не просто возможность изменения реакции и ее результата — выражения мутации (об этом мы уже говорили), а возможность изменения характера зависимости этого выражения от факторов внешней среды (рис. 26).
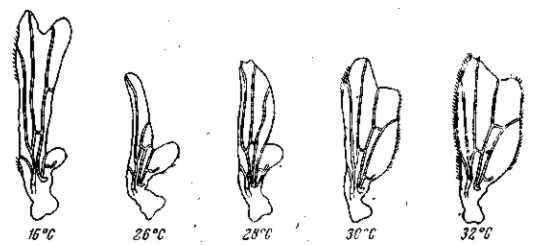
Рис. 24. Зависимость формы и величины крыльев гетерозиготы vestigial X pennant от температуры. Реакция меняет свое направление. При повышении температуры от 16 до 220 крылья уменьшаются; при 22–26° их средние размеры сохраняются постоянными; при дальнейшем повышении температуры крылья вновь увеличиваются. По М- Г. и М. JI. Хари, 1936.
В этом направлении был поставлен ряд интересных экспериментальных исследований М. Камшиловым, в особенности над мутацией безглазия (eyeless) дрозофилы. Из результатов этих исследований в данной связи очень большое значение имеет экспериментальное доказательство возможности полного извращения реакции. Нормально мутация безглазия получает наиболее типичное выражение при культивировании на влажном субстрате. Путем искусственного отбора на максимальное выражение редукции глаза в культуре на подсохшем корме удалось добиться того, что мухи, выводившиеся на влажном корме, обладали большим числом фасеток глаза, чем мухи, выводившиеся на подсохшем субстрате. Если в исходной линии уменьшение влажности связано с увеличением размеров глаза (т. е. уменьшением выражения мутации), то в экспериментально полученной линии уменьшение влажности ведет к уменьшению числа фасеток (усиление выражения мутации). В результате искусственного отбора получена, следовательно, линия, обладающая иной зависимостью морфо-генетического процесса (развития глаза) от внешнего фактора (влажности субстрата, чем исходная форма. Интересно, что имеющаяся зависимость может быть не только изменена, но и уничтожена. При чередующемся отборе по тому же выражению безглазия на влажном и на подсохшем субстрате удалось добиться такого наследственного изменения линии, что эта мутация получала в обоих случаях одинаковое выражение. Признак, зависящий от внешнего фактора (влажности субстрата), превратился в независимый. Характерная форма реагирования мутации eyeless оказалась погашенной через искусственный отбор малых наследственных изменений.
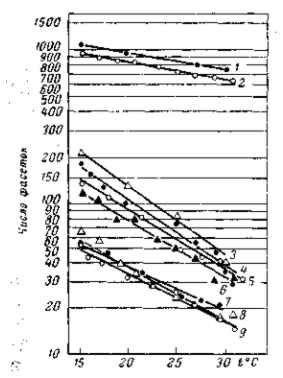
Рис. 25. Зависимость числа фасеток глаза нормальной дрозофилы и гомозиготных мутаций Ваг и Ultra-Bar от температуры. 1 — нормальная «дикая» форма; 2 — мутация, обратная к норме (получена в серии Ваг); 3 —культура Ваг, не подвергавшаяся отбору; 4–6 — культуры Ваг, подвергавшиеся легкому отбору на уменьшение числа фасеток; 7–9 — разные линии Ultra-Bar. Температурная реакция этих мутаций имеет такой же характер, как и у нормальных мух, но выражена гораздо резче (норма более стабильна): если у нормы при повышении температуры от 15 до 30° число фасеток падает от 1000 до 800, то у Ваг оно при тех же условиях падает примерно от 200 до 40 (т. е. в 5 раз). По А. Г. Гершу, 1934
К вопросу о значении этого обстоятельства мы еще вернемся в следующем разделе (4). Здесь же отметим, что формы реагирования мутаций на изменения в факторах внешней среды могут быть в процессе отбора преобразованы. Следовательно, вредные выражения таких зависимостей могут быть либо погашены, либо видоизменены и в результате получат иное, быть может, благоприятное выражение.
Исследования М. Камшилова показали, кроме того, возможность установления зависимостей иного рода между развивающимся организмом и внешней средой. В опытах охлаждения четырехдневной личинки до 0° в продолжение 24 часов наблюдалась довольно значительная смертность. Повторение этого опыта в течение целого ряда поколений привело через естественный отбор к созданию линии дрозофилы, весьма устойчивой по отношению к такому охлаждению. Однако при этом был достигнут еще и следующий результат: в этой новой «холодоустойчивой» линии «приспособление» к охлаждению достигло такого уровня, что ее жизнеспособность при непродолжительном охлаждении личинки (в четырехдневном возрасте) оказывалась даже более высокой, чем без охлаждения.
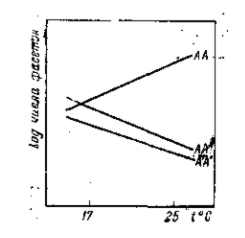
Рис. 26. Температурная зависимость числа фасеток глаза у гомозиготных самок (А'А') и Infra-Bar (AA) и у соответствующей гетерозиготы (АА'). Для графика применены логарифмы числа фасеток. Обе мутации дают прямо противоположную реакцию на изменение температуры. Однако у гетерозиготы не только не происходит взаимного погашения противоположных выражений этой зависимости, но характерная для Ваг обратная зависимость от температуры выражена даже еще резче, чем у гомозиготы. По А. Г. Гершу, 1930
Это показывает, что естественный отбор особей, развивающихся в определенных условиях внешней среды, приводит к установлению новых зависимостей между развивающимся организмом и факторами внешней среды. В некоторых случаях типичной смены факторов эта смена становится более или менее необходимым условием нормального развития организма. Вместе с тем и неблагоприятный фактор внешней среды может получить положительное значение в развитии организма (Камшилов, 1939а, б, 1941).
Все это подчеркивает возможность довольно быстрой перестройки зависимостей между развивающимся организмом и факторами внешней среды и показывает лишний раз, что реакции ор ганизма на изменения во внешних факторах имеют историческую базу. Благоприятные формы реагирования на внешние факторы не даны как изначальные свойства организма, а являются результатом естественного отбора особей в конкретных условиях развития организма в определенной внешней среде. Новые, элементарные формы реагирования любой новой мутации (как часть ее нормы реакций) обычно не имеют положительного значения. Однако в процессе естественного отбора (малых мутаций — модификаторов) эти формы реагирования могут менять свойхарактери быть либо погашенными, либо преобразованными в благоприятные, т. е. адаптивные, формы морфоге-нетических реакций.
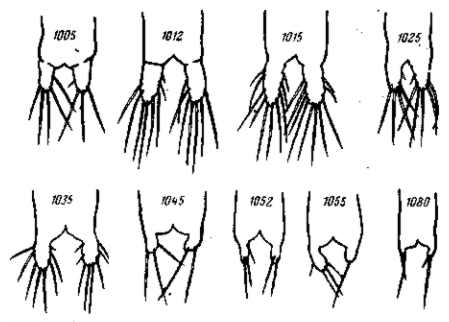
Рис. 27. Модификация Artemia salina в солевых растворах разной концентрации (на рисунке цифрами обозначены удельные веса). Культура проходит при уд. весе 1015 в явно оптимальных условиях — развитие фурк и щетинок достигает максимальной полноты. В наиболее концентрированных растворах фурки и щетинки совсем не развиваются. По A. Abonyi, 1915
б. Явление недоразвития в ненормальных условиях. На грани морфозов и адаптивных модификаций стоят явления недоразвития, нередко наблюдаемые в тех случаях, когда организм попадает в ненормальные условия внешней среды.
Мы только что упомянули, что в течение исторического развития организма (иногда даже в течение сравнительно кратковременного эксперимента) в определенной внешней среде, в особенности при наличии известной смены факторов, устанавливаются нередко зависимости такого рода, что некоторая последовательность в смене внешних факторов становится обязательным условием для нормального развития организма.
Такие зависимости весьма обычны для организмов с точно определенным, сезонным циклом размножения. Яйца многих насекомых, а также мелких ракообразных требуют зимнего охлаждения для начала их развития. То же самое относится иногда и к другим стадиям развития, —иначе развитие не возобновляется после «диапаузы». Особенно это характерно для растений, требующих для полного развития и созревания известного предварительного охлаждения для прорастания и весеннего пробуждения, а также определенных доз света, притом в виде светового дня соответствующей длительности. Отсутствие во внешней среде необходимых условий для индивидуального развития приводит такие организмы к явлениям недоразвития. Во многих случаях это означает отсутствие цветения у растения в чуждом климате или отсутствие половою созревания у животного. Организм не может противостоять таким изменениям и в новой среде не размножается. В других случаях явления недоразвития выражены не столь резко. Организм размножается, но оказывается соответственно модифицированным. Такие модификации нельзя назвать адаптациями, хотя они и сопутствуют переживанию организма в измененных условиях внешней среды.
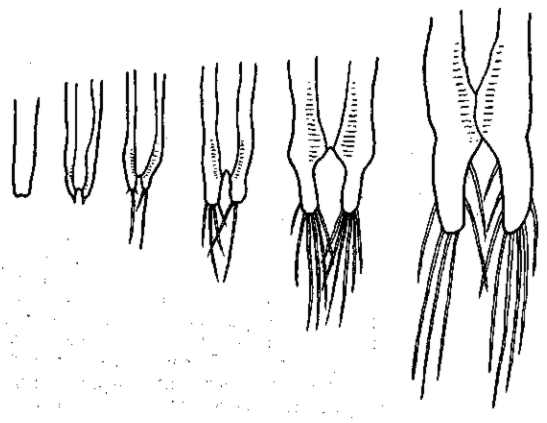
Рис. 28 Стадии индивидуального развития фурк и щетинок у Artemia salina в оз. Майли-Сор (Сев. Казахстан) при содержании С1 в воде около 6: 100 и уд. весе около 1,10. По О. И. Шмальгаузен
Так, например, многие сухопутные растения, попадая в воду, не гибнут в ней, а продолжают расти. Они, однако, модифицируются, и такие изменения имеют характер общего недоразвития (lingerer). Соленоводный жаброног Artemia salina может жить и развиваться в очень соленой воде, приближающейся к насыщению. Однако он модифицируется в ней, изменяясь именно в сторону общего недоразвития — размеры рачка значительно уменьшаются, последние членики брюшка и фурка сохраняют личиночное строение (рис, 27, 28). Половое созревание можно считать преждевременным и всю эту ультрагалинную модификацию рассматривать как частный случай неотении (Abonyi, 1915). Морская мизида Mysis oculata, попадающая у нас на севере в условия опресненной воды, уменьшается в размерах и приближается к пресноводной Mysis relicta. Такое же измельчание наблюдается и при переходе морского таракана Chiridothea sibirica в устья рек и в пресные озера. Пресноводные модификации С. entemon-C. vetterensis отличаются уменьшенными размерами и узким тельзоном. Двустворчатый моллюск — мидия Mytilus edulis — значительно уменьшается в размерах, переходя в опресненные воды (например, Ботнического залива). То же самое касается многих других морских животных, которые при переходе в пресные воды приобретают карликовые размеры и недоразвитые формы.
Б. АДАПТИВНЫЕ МОДИФИКАЦИИ
В предыдущем разделе мы рассмотрели элементарные реакции организма, попадающего в совершенно новые условия внешних факторов, или реакции мутантного организма, которые также связаны с совершенно новыми взаимоотношениями между организмом и факторами внешней среды. Теперь мы познакомимся с уже установившимися в предшествующей истории формами реагирования. Некоторые из них имеют общефизиологический характер. В процессе эволюции они лишь вводятся в известные пределы оптимальных изменений. Таковы изменения скорости роста и общей величины тела, плодовитости, вегетации и цветения у растений в зависимости от питания, условий влажности и температуры. Другие реакции имеют более специальный (но в основе, конечно, также физиологический) характер, выражаясь в специфических приспособлениях организма и его органов к известным изменениям в физических факторах внешней среды. Мы их называем физиоген-ными модификациями. Например, изменения поверхности листьев в зависимости от интенсивности освещения, изменения пигментации у животных или адаптивные реакции в органах терморегуляции при изменении температуры. Наконец, третьи реакции выражаются в функциональном приспособлении органов и представляют, следовательно, лишь косвенный ответ организма на изменения в факторах внешней среды. Эта последняя категория функциональных модификаций может лишь условно быть отграничена от физиогенных модификаций, причем мы здесь отмечаем лишь относительно большую активность самого организма (рис. 29). АПТИВНЫХ РЕАКЦИЙ 291
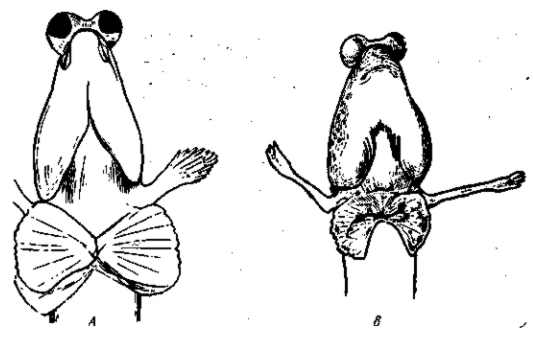
Рис. 29. Модификация прибрежной рыбы Periophthalmus variabilis А — нормальная водная форма; В — наземная модификация, полученная в эксперименте применением тироксина. По I. W. Harms, 1934
а. Модификации общефизиологического характера. Естественно, что возможности роста ограничиваются имеющимися запасами питательных материалов (у зародыша) и их дальнейшим поступлением при самостоятельном питании развивающегося организма. Точно так же понятно, что и верхний предел скорости роста ограничивается нередко возможной интенсивностью питания. Однако в процессе эволюции устанавливаются известные оптимальные нормы роста, определяемые экологическим положением организма и, в особенности, его соотношениями с другими организмами. Модификации возможны тогда лишь в сравнительно узких пределах этой нормы. В качестве приспособления мы должны здесь рассматривать не самую модификацию, а ее ограничение известными пределами. То же самое касается и скорости роста организма, которая имеет свой максимум. Минимум скорости роста находится в непосредственной зависимости от питания. Одаако сниженная скорость роста может быть компенсирована при позднейшем усилении питания. В этих явлениях компенсации мы также видим приспособление организма к восстановлению нормальных размеров при их нарушении вследствие временной недостачи питательных материалов. Плодовитость организма таюке зависит от питания. Однако уровень плодовитости определяется и экологическими условиями, т. е. положением организма в условиях его борьбы за существование. Адаптивной является и в этом случае не сама плодовитость и ее модификации, а именно ее ограничение известным оптимальным для данного вида уровнем.
Общая поверхность листьев растения и развитие его корневой системы подвержены также значительным модификациям, определяемым освещением и влажностью. В основе этих модификаций лежат также явления роста, определяемые условиями питания (ассимиляции). Адаптивный характер имеет опять-таки ограничение роста оптимальными в данных условиях пределами.
Во всех этих случаях мы имеем дело с общефизиологическими процессами, которые оказываются всегда очень изменчивыми вследствие постоянного обилия малых «физиологических» мутаций, выражающихся в изменении интенсивности обмена веществ, скорости роста, его длительности, а следовательно, и в изменении нормальных его пределов. То же самое касается и плодовитости, ее зависимости от питания и ее нормального и предельного выражения. Все эти зависимости явно меняются и в отдельных мутациях, и в процессе эволюции, определяемом естественным отбором оптимальных форм реагирования и пределов роста, оптимальной плодовитости и ее возможных пределов при разных условиях питания (температуры, влажности, освещения и т. п.).
б. Физиогенные модификации. Яснее выражается адаптивный характер модификаций, связанных более непосредственно с изменениями в физических факторах внешней среды. Так, например, в зависимости от условий освещения и влажности меняется величина, а нередко и форма листовой пластинки растений. Развиваются световые и теневые формы листьев, которые обладают оптимальной структурой для ассимиляции на прямом солнечном свету или в тени и вместе с тем соответствуют различным условиям транспирации. Необходимость ее ограничения при недостатке влажности ведет к дальнейшим адаптивным преобразованиям световых листьев в ксерофитные. Световые листья не только мельче и, как правило, менее рассечены, но обладают и более толстой кутикулой и узкими межклетниками, а также частыми, но более мелкими устьицами. Ксерофитные листья обладают и другими приспособлениями для удержания воды в тканях и для защиты от чрезмерной инсоляции (например, опушение листьев). Все эти приспособления весьма изменчивы. У некоторых ксерофитов прилистники или часть листьев преобразуются в колючки, но при развитии в условиях достаточной влажности вместо колючек вновь развиваются листья.
Во всех этих случаях модификации осуществляются через зависимость процессов роста и формообразования от условий освещения и степени влажности. У растений процессы роста сильно; зависят от вакуолизации растущих клеток: поэтому обеспеченность водой и ограничение испарения естественно способствуют более свободному росту растения в тени. С другой стороны, недостаток влаги в наземных частях и обильное испарение на солнечном свету ограничивают этот рост. Таким образом, адаптивные модификации здесь осуществляются на базе таких же общефизиологических процессов, как уже рассмотренные. В данных случаях они имеют лишь более локализованный характер и дополняются зависимыми же формообразовательными реакциями, основанными в значительной мере на неравномерном росте частей и на изменениях в гистологической дифференцировке. Формообразование; у растений вообще весьма зависимо от физических факторов,-а мутации, выражающиеся в изменении формы листьев и степени их расчлененности, принадлежат к наиболее обычным мутациям растительных организмов. Фенотипическая изменчивость таких мутаций и лежит, очевидно, в основе исторического развития способности к указанным адаптивным модификациям. У животных некоторым аналогом таких зависимостей могут служить температурные реакции млекопитающих, выражающиеся в изменении величины поверхности тела, его придатков и в изменении шерстного покрова. Эти реакции основываются также на широко распространенных физиологических процессах, связанных с общими закономерностями роста. Высокая температура способствует ускорению клеточных делений, а следовательно, повышению скорости роста при одновременном, однако, уменьшении размеров клеток, ускорении гистологической их дифференцировки, сокра^ щении дефинитивных размеров тела и увеличении его относительной поверхности. Об этом свидетельствует целый ряд экспериментальных данных по личинкам амфибий, эмбрионам птиц.
Мутации и расовые различия, сопровождающиеся изменением температурных реакций (пигментообразование) в волосах горностаевого кролика, по Ильину (см. рис. 8), вообще говоря, известны так же, как известны и наследственные изменения размеров тела,, размеров конечностей и придатков и, в особенности, длины, толщины, числа волос и формы их роста. Однако мутационные изменения зависимости роста этих частей от температуры, по-видимому, не изучены.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 469; Нарушение авторских прав?; Мы поможем в написании вашей работы!