КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
В. Аллостерическая регуляция аэробного распада глюкозы и глюкогенеза в печени энергетическим статусом клетки 2 страница
В. Метаболизм галактозы
Галактоза образуется в кишечнике в результате гидролиза лактозы. Чтобы превратить галактозу в глюкозу, необходимо изменить оптическую конфигурацию Н- и ОН-групп С4 атома в галактозе, т.е. провести реакцию эпимеризации. Эта реакция в клетке возможна только с УДФ-произ-водным галактозы. УДФ-галактоза образуется из УДФ-глюкозы (метаболит в синтезе гликогена) в ходе реакции, катализируемой уридилфосфат-4-эпимеразой (рис. 7-70, 7-71).
Однако включению галактозы в описанную реакцию эпимеризации предшествует её фос-форилирование с образованием галактозо-1-фосфата (реакция 1 на рис. 7-70). Далее галактозо-1-фосфат замещает остаток глюкозы в УДФ-глюкозе с образованием УДФ-галактозы (реакция 2), т.е. прямая реакция фосфорилированной галактозы с УТФ не происходит.
Реакцию 2 можно рассматривать как перенос уридильного остатка с УДФ-глюкозы на галактозу, поэтому фермент назван галактозо-1 -фосфатуридилтрансферазой (ГАЛТ).
Затем галактоза в составе нуклеотида включается в реакцию эпимеризации, в которой участвует эпимераза - NAD-зависимый фермент, катализирующий окисление и восстановление галактозы по С4 углеродному атому (реакция 3).
Эпимераза может работать и в другом направлении, преобразуя УДФ-глюкозу в УДФ-галактозу. Эта обратная эпимеризация важна для синтеза галактозильных остатков в гликолипидах и гликопротеинах. Кроме того, галактоза необходима для синтеза лактозы в грудных железах. В период лактации галактоза не является незаменимым компонентом пищи, так как может образовываться из глюкозы.
Глюкозо-1-фосфат, образованный в реакции 2, может включаться в разные метаболические пути: 1) синтез гликогена после реакции с УДФ и образования УДФ-глюкозы; 2) превращение в печени
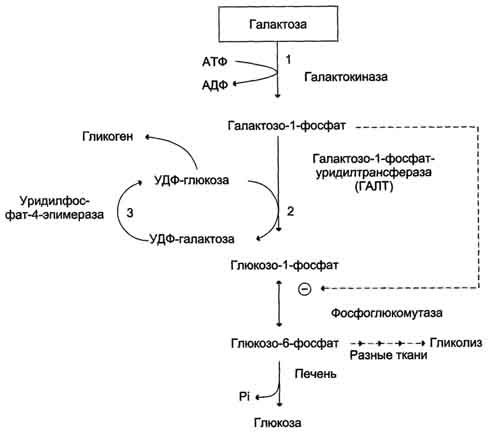
Рис. 7-70. Обмен галактозы.
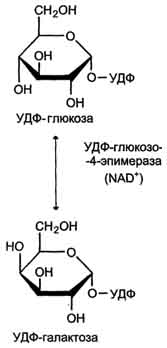
Рис. 7-71. Реакция эпимеризации УДФ-глюкозы в УДФ-галактозу.
в свободную глюкозу и поддержание её концентрации в крови; 3) катаболизм, сопряжённый с синтезом АТФ, и т.д. (см. рис. 7-70).
Г. Нарушения метаболизма галактозы
Обмен галактозы особенно интересен в связи с наследственным заболеванием - галактоземией.
Галактоземия возникает при нарушении обмена галактозы, обусловленном наследственным дефектом любого из трёх ферментов, включающих галактозу в метаболизм глюкозы (табл. 7-6).
Галактоземия, вызванная недостаточностью галактозо-1-фосфатуридилтрансферазы (ГАЛТ), наиболее хорошо изучена. Это заболевание проявляется очень рано, и особенно опасно для детей, так как основным источником углеводов для них служит материнское молоко, содержащее лактозу. Ранние симптомы дефекта ГАЛТ: рвота, диарея, дегидратация, уменьшение массы тела, желтуха. Они появляются вскоре после рождения, как только ребёнок начинает получать молоко. В крови, моче и тканях повышается концентрация галактозы и галактозо-1-фосфата. В тканях глаза (в хрусталике) галактоза восстанавливается альдоредуктазой с образованием галактитола (дульцита). В этой реакции в качестве донора водорода используется
NADPH. Восстановление галактозы происходит и в ходе нормального метаболизма, но протекает с небольшой скоростью. При галактоземии галактитол накапливается в стекловидном теле и связывает большое количество воды. Вследствие этого нарушается баланс электролитов, а чрезмерная гидратация хрусталика приводит к развитию катаракты, которая наблюдается уже через несколько дней после рождения.
Тяжёлые последствия дефекта ГАЛТ наблюдают в печени. Это связано с накоплением галактозо-1-фосфата и его токсическим действием на гепатоциты. В результате возникают нарушения функции печени: гепатомегалия, жировая дистрофия. В почках таких больных также повышена концентрация галактитола и галактозо-1-фосфата, что влияет на их функции. Отмечают нарушения в клетках полушарий головного мозга и мозжечка, в тяжёлых случаях - отёк мозга, задержку умственного развития, возможен летальный исход.
Для галактоземии, вызванной дефектом галактокиназы, тоже характерна катаракта, но при этом заболевании, в отличие от дефекта ГАЛТ, не отмечают нарушений функций печени, почек, мозга. Наиболее тяжёлые последствия снижения активности ГАЛТ связывают с влиянием галактозо-1-фосфата на активность других ферментов, участвующих в углеводном
Таблица 7-6. Нарушения обмена галактозы
| Дефектный фермент (частота) | Блокируемая реакция | Клинические проявления и лабораторные данные |
| Галактокиназа (1:500 000) | Галактоза + АТФ → Галактозо-1-фосфат + АДФ | Галактоземия, галактозурия, катаракта. Активность фермента в эритроцитах нормальная. |
| Галактозо-1-фосфатуридилтрансфераза (1:40000) | Галактозо-1-фосфат + УДФ-глюкоза → УДФ-галактоза + Глюкозо-1-фосфат | Галактоземия, галактозурия, галактозо-1-фосфатемия, катаракта. Тенденция к гипогликемии, компенсаторная мобилизация жиров, цирроз печени, нарушения функции почек. Гепатомегалия, задержка психического развития. Активность фермента в эритроцитах снижена. |
| Уридилфосфат-4-эпимераза (1:1000000) | УДФ-глюкоза ↔ УДФ-галактоза | Галактоземия, галактозурия. Тяжёлых клинических проявлений нет. Описаны единичные случаи заболевания. |
Таблица 7-7. Некоторые варианты генетического дефекта ГАЛТ
| Изменения в структуре ГАЛТ | Проявления |
| Асн→Асп | Признак Дюарта. У гетерозигот при этом варианте активность фермента составляет 75% от нормальной. Гомозиготный фенотип Дюарта обычно связан с 50% потерей активности. Пациенты с синдромом Дюарта могут быть здоровыми, несмотря на структурную аномалию ГАЛТ. |
| Глн→Арг | Проявляется как тяжёлая галактоземия. Причина - мутация типа замены нуклеотида 591 в гене фермента. Активность ГАЛТ составляет 10% от нормы. Эта форма встречается в 70% случаев заболевания галактоземией среди европеоидов, частота - 1:338 886. |
| Сер→Лей | Заболевание описано у чернокожих пациентов и названо «чёрный признак». Галактоземия проявляется как результат недостаточной активности ГАЛТ в печени и эритроцитах. Активность ГАЛТ в печени составляет 10% от нормы. Тем не менее отмечалась утилизация некоторого количества галактозы, что объяснялось развитием альтернативного пути. Причина - мутация типа замены 1 158-го нуклеотида в гене фермента. |
| Арг→Три | Тяжёлая форма галактоземии. Причина - миссенс-мутация нуклеотида 1025 в гене фермента. Активность ГАЛТ отсутствует. |
| Лиз→Асн | Широко распространённая мутация при галактоземии. |
обмене (фосфоглюкомутазы, глюкозо-6-фосфатдегидрогеназы).
Известно несколько форм галактоземии, причиной которой является недостаточность ГАЛТ (табл. 7-7).
Некоторые дефекты в строении ГАЛТ приводят лишь к частичной потере активности фермента. Поскольку в норме ГАЛТ присутствует в организме в избытке, то снижение его активности до 50%, а иногда и ниже может клинически не проявляться.
При диагностике галактоземии исследуют мочу на содержание галактозы, собранную после нескольких кормлений молоком. При обнаружении у ребёнка катаракты его обследуют на недостаточность галактокиназы и ГАЛТ. Наличие галактозы в моче при отсутствии нарушений функции печени указывает на дефект галактокиназы. При обследовании проведение теста с нагрузкой галактозой не рекомендуется, так как этот тест опасен для больных. Лечение заключается в удалении галактозы из рациона.
364:: 365:: 366:: 367:: 368:: 369:: Содержание
297:: Содержание

ОБМЕН УГЛЕВОДОВ
Углеводы входят в состав живых организмов и вместе с белками, липидами и нуклеиновыми кислотами определяют специфичность их строения и функционирования. К углеводам относят соединения, обладающие разнообразными и зачастую сильно отличающимися функциями. Углеводы участвуют во многих метаболических процессах, но прежде всего они являются основными поставщиками энергии. На долю углеводов приходится примерно 75% массы пищевого суточного рациона и более 50% от суточного количества необходимых калорий. Однако неправильно сводить функцию углеводов только к энергетическому обеспечению процессов жизнедеятельности организма. Следует отметить и структурную роль углеводов. Так, в виде гликозаминогликанов углеводы входят в состав межклеточного матрикса. Большое число белков (ферменты, белки-транспортёры, белки-рецепторы, гормоны) - гликопротеины, углеводная составляющая которых повышает их специфичность. Например, различия в строении олигосахаридных фрагментов клеточной оболочки эритроцитов обеспечивают групповую принадлежность крови. Из углеводов в процессе метаболизма образуется большое число органических соединений, которые служат исходными субстратами для синтеза липидов, аминокислот, нуклеотидов. Производные углеводов - глюкурониды - участвуют в детоксикации ксенобиотиков и инактивации веществ эндогенного происхождения. Углеводы могут быть синтезированы в организме с использованием других метаболитов: некоторых аминокислот, глицерина, молочной кислоты. Углеводы нельзя считать незаменимыми компонентами пищи. Однако если исключить углеводы из диеты, то следствием может быть гипогликемия, для компенсации которой будут расходоваться белки и липиды. Таким образом, углеводы - обязательные пищевые компоненты, потому что помимо их основной энергетической функции (клеточные "дрова") углеводы участвуют во многих метаболических клеточных процессах.
297:: Содержание
298:: 299:: 300:: 301:: 302:: 303:: 304:: 305:: Содержание
I. СТРОЕНИЕ УГЛЕВОДОВ
Термин "углеводы", предложенный в XIX столетии, был основан на предположении, что все углеводы содержат 2 компонента - углерод и воду, и их элементарный состав можно выразить общей формулой Cm(H2O)n. Хотя из этого правила есть исключения и оно не абсолютно точно, тем не менее указанное определение позволяет наиболее просто характеризовать класс углеводов в целом. К тому же попытка, предпринятая Комиссией по химической номенклатуре, заменить термин "углеводы" на "глициды" не удалась. Новый термин не получил широкого признания. Термин "углеводы" укоренился и общепризнан.
Углеводы можно разделить на 3 основные группы в зависимости от количества составляющих их мономеров: моносахариды, олигосахариды и полисахариды.
А. Моносахариды
Моносахариды - производные многоатомных спиртов, содержащие карбонильную группу. В зависимости от положения в молекуле карбонильной группы моносахариды подразделяют на альдозы и кетозы.
Альдозы содержат функциональную альдегидную группу -НС=О, тогда как кетозы содержат кетонную группу >С=О. Название моносахарида зависит от числа составляющих его углеродных атомов, например альдотриозы, кетотриозы, альдогексозы, кетогексозы и т.д.
Моносахариды по строению можно отнести к простым углеводам, так как они не гидролизуются при переваривании, в отличие от сложных, которые при гидролизе распадаются с образованием простых углеводов. Строение основных представителей моносахаридов показано на рис. 7-1.
В пище человека (фрукты, мёд, соки) содержится небольшое количество моносахаридов, в основном глюкоза и фруктоза.
Глюкоза является альдогексозой. Она может существовать в линейной и циклической формах. Циклическая форма глюкозы, предпочтительная в термодинамическом отношении, обусловливает химические свойства глюкозы. Как и все гексозы, глюкоза имеет 4 асимметричных углеродных атома, обусловливающих наличие сте-реоизомеров. Возможно образование 16 стереоизомеров, наиболее важные из которых D- и L-глюкоза. Эти типы изомеров зеркально отображают друг друга (рис. 7-2).
Расположение Н- и ОН-групп относительно пятого углеродного атома определяет принадлежность глюкозы к D- или L-ряду. В организме млекопитающих моносахариды находятся в D-конфигурации, так как к этой форме глюкозы специфичны ферменты, катализирующие её превращения. В растворе при образовании циклической формы моносахарида образуются ещё 2 изомера (α- и β-изомеры), называемые аномерами, обозначающие определённую конформа-цию Н- и ОН-групп относительно С, (рис. 7-3). У α-D-глюкозы ОН-группа располагается ниже плоскости кольца, а у β-D-глюкозы, наоборот, над плоскостью кольца.
Фруктоза является кетогексозой (кетогругша находится у второго углеродного атома). Фруктоза так же, как и глюкоза, существует в циклической форме, образуя α- и β-аномеры (рис. 7-4).
Б. Реакции моносахаридов
Присутствие гидроксильных, альдегидных и кетонных групп позволяет моносахаридам вступать в реакции, характерные для спиртов, альдегидов или кетонов. Эти реакции довольно многочисленны. В данном разделе будут описаны лишь некоторые из них, причём в основном имеющие наибольшее биологическое значение.
В этом разделе основные реакции моносахаридов рассмотрены на примере D-глюкозы (рис. 7-5), хотя надо иметь в виду, что в метаболизме углеводов принимают участие и другие моносахариды, а также их производные.
Мутаротация, или аномеризация -взаимопревращение аномерных форм моносахаридов, α- и β-формы аномеров находятся в растворе в состоянии равновесия. При достижении этого равновесия происходит мутаротация - размыкание и замыкание пиранового кольца и, соответственно, изменение расположения Н- и ОН-групп при первом углероде моносахарида.
Образование гликозидов. Гликозидная связь имеет важное биологическое значение, потому что именно с помощью этой связи осуществляется ковалентное связывание моносахаридов в составе олиго- и полисахаридов. При образовании гликозидной связи аномерная ОН-группа одного моносахарида взаимодействует с ОН-группой другого
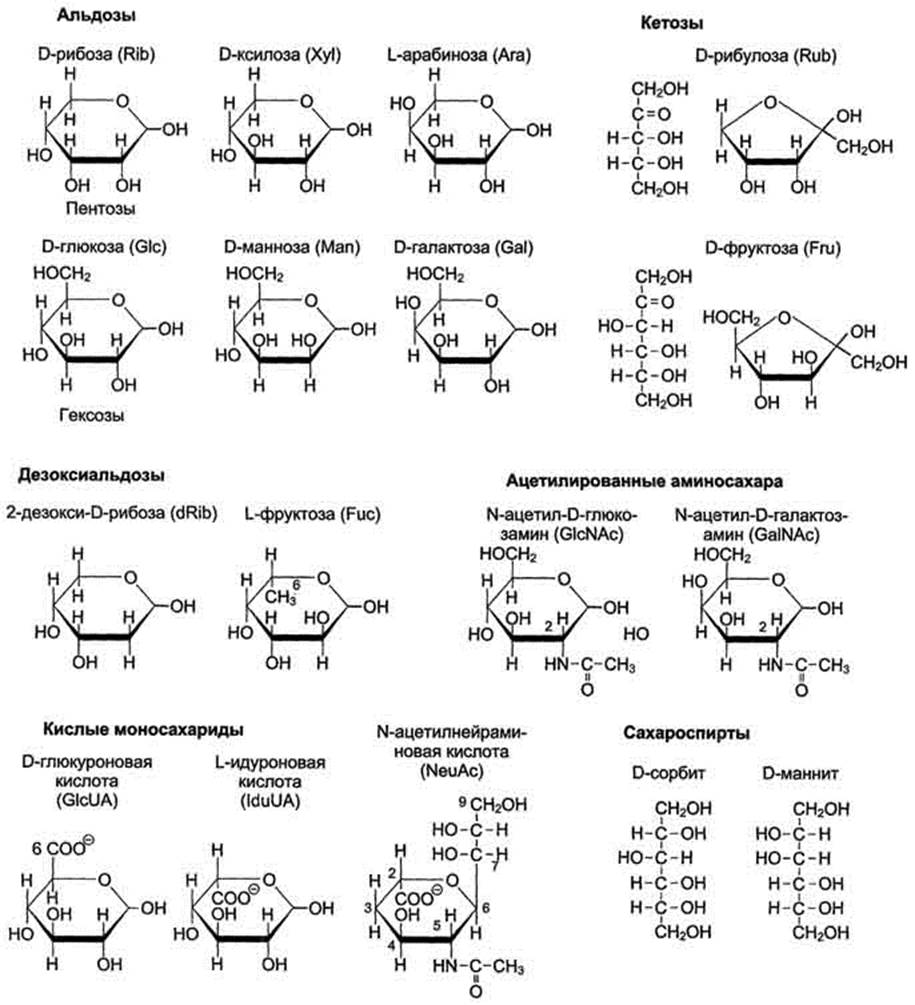
Рис. 7-1. Важнейшие моносахариды.
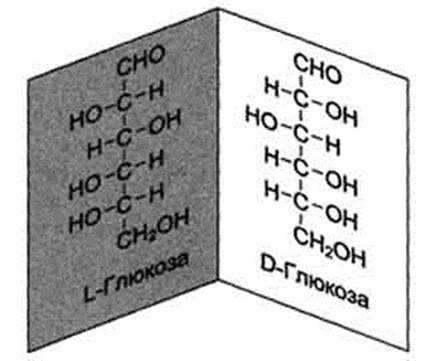
Рис. 7-2. D- и L-изомеры глюкозы.
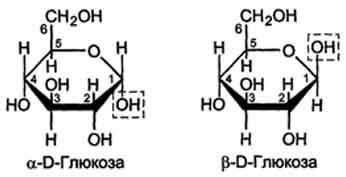
Рис. 7-3. α- и β -аномеры D-глюкозы.
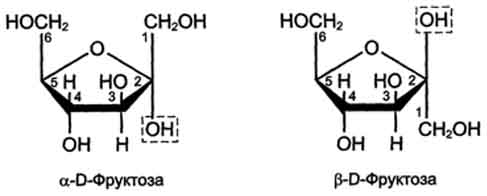
Рис. 7-4. α- и β -аномеры D-фруктозы.
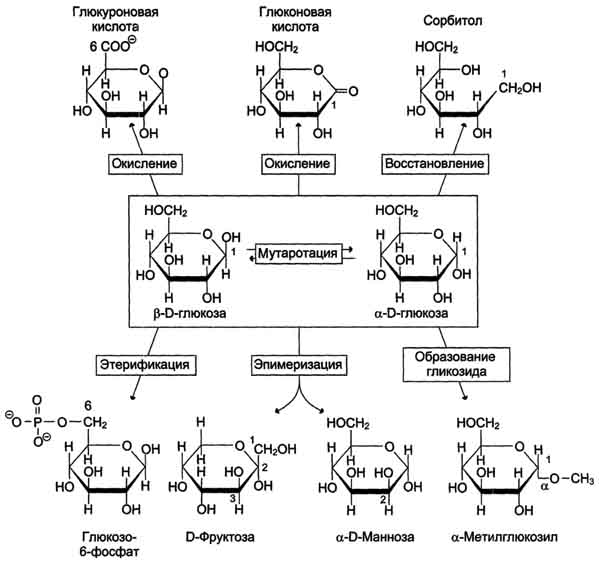
Рис. 7-5. Реакции моносахаридов.
моносахарида или спирта. При этом происходят отщепление молекулы воды и образование О-гликозидной связи. Все линейные олигомеры (кроме дисахаридов) или полимеры содержат мономерные остатки, участвующие в образовании двух гликозидных связей, кроме концевых остатков, образующих только одну гликозидную связь. Некоторые гликозидные остатки могут образовывать три гликозидные связи, что характерно для разветвлённых олиго- и полисахаридов. Олиго- и полисахариды могут иметь концевой остаток моносахарида со свободной аномерной ОН-группой, не использованной при образовании гликозидной связи. В этом случае при размыкании цикла возможно образование свободной карбонильной группы, способной окисляться. Такие олиго- и полисахариды обладают восстанавливающими свойствами и поэтому называются восстанавливающими, или редуцирующими (рис. 7-6).
Аномерная ОН-группа моносахарида может взаимодействовать с NН2-группой других соединений, что приводит к образованию N-гликозидной связи. Подобная связь присутствует в нуклеотидах и гликопротеинах (рис. 7-7).
Этерификация. Это реакция образования эфирной связи между ОН-группами моносахаридов и различными кислотами. В метаболизме углеводов важную роль играют фосфоэфиры - эфиры моносахаридов и фосфорной кислоты. В метаболизме глюкозы особое место
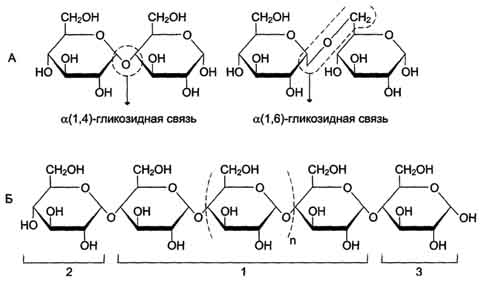
Рис. 7-6. Строение полисахарида. A. Образование α-1,4- и α-1,6-гликозидных связей. Б. Строение линейного полисахарида: 1 - α-1,4-гликозидные связи между мономерами; 2 - невосстанавливающий конец (невозможно образование свободной карбонильной группы у аномерного углерода); 3 - восстанавливающий конец (возможно размыкание цикла с образованием свободной карбонильной группы у аномерного углерода).
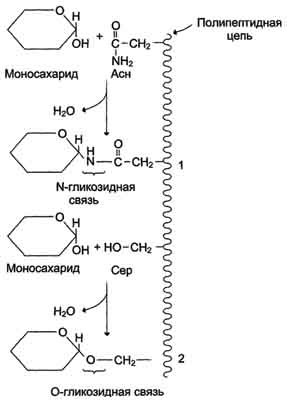
Рис. 7-7. Образование О- и N-гликозидных связей в гликопротеинах. 1 - N-гликозидная связь между амидной группой аспарагина и ОН-группой моносахарида; 2 - О-гликозидная связь между ОН-группой серина и ОН-группой моносахарида.
занимает глюкозо-6-фосфат. Образование глюкозо-6-фосфата происходит в ходе АТФ-зависимой реакции при участии ферментов, относящихся к группе киназ. АТФ в данной реакции выступает как донор фосфатной группы. Фосфоэфиры моносахаридов могут образовываться и без использования АТФ. Например, глюкозо-1-фосфат образуется из гликогена при участии Н3РО4. Физиологическое значение фос-фоэфиров моносахаридов заключается в том, что они представляют собой метаболически активные структуры. Реакция фосфорилирования моносахаридов важна для метаболизма ещё и потому, что клеточная мембрана мало проницаема для этих соединений, т.е. клетка удерживает моносахариды благодаря тому, что они находятся в фосфорилированной форме.
Окисление и восстановление. При окислении концевых групп глюкозы -СНО и -СН2ОН образуются 3 различных производных. При окислении группы -СНО образуется глюконовая кислота. Если окислению подвергается концевая группа -СН2ОН, образуется глюкуроновая кислота. А если окисляются обе концевые группы, то образуется сахарная кислота, содержащая 2 карбоксильные группы. Восстановление первого углерода приводит к образованию сахароспир-та - сорбитола.
В. Олигосахариды
Олигосахариды содержат несколько (от двух до десяти) остатков моносахаридов, соединённых гликозидной связью. Дисахариды - наиболее распространённые олигомерные углеводы, встречающиеся в свободной форме, т.е. не связанной с другими соединениями. По химической природе дисахариды представляют собой гликозиды, которые содержат 2 моносахарида, соединённые гликозидной связью в α- или β-конфигурации. В пище содержатся в основном такие дисахариды, как сахароза, лактоза и мальтоза (рис. 7-8).
Сахароза - дисахарид, состоящий из α-D-глюкозы и β-D-фруктозы, соединённых α,β-1,2-гликозидной связью. В сахарозе обе аномерные ОН-группы остатков глюкозы и фруктозы участвуют в образовании гликозидной связи. Следовательно, сахароза не относится к восстанавливающим сахарам. Сахароза - растворимый дисахарид со сладким вкусом. Источником сахарозы служат растения, особенно сахарная свёкла, сахарный тростник. Последнее объясняет возникновение тривиального названия сахарозы - "тростниковый сахар".
Лактоза - молочный сахар; важнейший дисахарид молока млекопитающих. В коровьем молоке содержится до 5% лактозы, в женском молоке - до 8%. В лактозе аномерная ОН-группа первого углеродного атома остатка D-галактозы связана β-гликозидной связью с четвёртым углеродным атомом D-глюкозы (β-1,4-связь). Поскольку аномерный атом углерода остатка глюкозы не участвует в образовании гликозидной связи, следовательно, лактоза относится к восстанавливающим сахарам.
Мальтоза поступает с продуктами, содержащими частично гидролизованный крахмал, например, солод, пиво. Мальтоза также образуется при расщеплении крахмала в кишечнике. Мальтоза состоит из двух остатков D-глюкозы, соединённых α-1,4-гликозидной связью.
Изомальтоза - промежуточный продукт, образующийся при расщеплении крахмала в кишечнике. Состоит из двух остатков D-глюкозы, но соединены эти моносахариды α-1,6-гликозидной связью.
Г. Полисахариды
Структурные различия между полисахаридами определяются:
- строением моносахаридов, составляющих цепь;
- типом гликозидных связей, соединяющих мономеры в цепи;
- последовательностью остатков моносахаридов в цепи.
В зависимости от строения остатков моносахаридов полисахариды можно разделить на гомополисахариды (все мономеры идентичны) и гетерополисахариды (мономеры различны). Оба типа полисахаридов могут иметь как линейное расположение мономеров, так и разветвлённое.
В зависимости от выполняемых ими функций полисахариды можно разделить на 3 основные группы:
- резервные полисахариды, выполняющие энергетическую функцию. Эти полисахариды служат источником глюкозы, используемым организмом по мере необходимости. Резервная функция этих углеводов обеспечивается их полимерной природой. Полисахариды
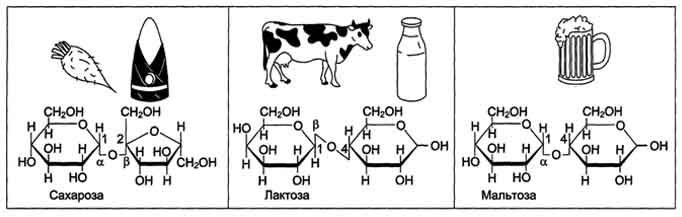
Рис. 7-8. Дисахариды пищи.
· менее растворимы, чем моносахариды, следовательно они не влияют на осмотическое давление и поэтому могут накапливаться в клетке, например, крахмал - в клетках растений, гликоген - в клетках животных;
- структурные полисахариды, обеспечивающие клеткам и органам механическую прочность (см. раздел 15);
- полисахариды, входящие в состав межклеточного матрикса, принимают участие в образовании тканей, а также в пролиферации и дифференцировке клеток. Полисахариды межклеточного матрикса водорастворимы и сильно гидратированы (см. раздел 15).
В пище человека в основном содержатся полисахариды растительного происхождения - крахмал, целлюлоза. В меньшем количестве поступает полисахарид животных - гликоген.
Крахмал - наиболее важный углеводный компонент пищевого рациона. Это резервный полисахарид растений, содержащийся в наибольшем количестве (до 45% от массы сухого вещества) в зёрнах злаков (пшеница, кукуруза, рис и др.), а также луковицах, стеблях и клубнях растений (в картофеле примерно 65%). Крахмал - разветвлённый полисахарид, состоящий из остатков глюкозы (гомогликан). Он находится в клетках растений в виде гранул, практически нерастворим в воде.
Крахмал состоит из амилозы и амилопектина (рис. 7-9). Амилоза - неразветвлённый полисахарид, включающий 200-300 остатков глюкозы, связанных α-1,4-гликозидной связью. Благодаря α-конфигурации глюкозного остатка, полисахаридная цепь имеет конформацию спирали. Синяя окраска при добавлении йода к раствору крахмала обусловлена наличием такой спирали. Амилопектин имеет разветвлённую структуру. В местах ветвления остатки глюкозы соединены α-1,6-гликозидными связями. Линейные участки содержат примерно 20-25 остатков глюкозы. При этом формируется древовидная структура, в которой имеется лишь одна аномерная ОН-группа. Крахмал - высокомолекулярное соединение, включающее сотни тысяч остатков глюкозы. Его молекулярная масса составляет порядка 105-108 Д.
Целлюлоза (клетчатка) - основной структурный полисахарид растений. Это самое распространённое органическое соединение на земле. Доля целлюлозы в клеточных стенках растений составляет 40-50%. Целлюлоза имеет молекулярную массу порядка 106 Д, длина молекулы может доходить до 6-8 мкм.
Целлюлоза - линейный полисахарид гомогликан, построенный из остатков глюкозы, соединённых между собой β-1,4-гликозидными связями. Пищеварительная система человека не имеет ферментов, гидролизующих β-связи в полисахаридах. Поэтому целлюлоза - неиспользуемый углевод, но этот пищевой компонент необходим для нормального протекания переваривания.
Гликоген - полисахарид животных и человека. Так же, как крахмал в растениях, гликоген в клетках животных выполняет резервную функцию, но, так как в пище содержится лишь небольшое количество гликогена, он не имеет пищевого значения.
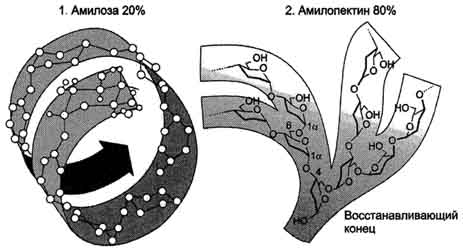
Рис. 7-9. Строение крахмала.
Гликоген представляет собой структурный аналог крахмала, но имеет большую степень ветвления: примерно на каждые 10 остатков глюкозы приходится одна α-1,6-гликозидная связь.
298:: 299:: 300:: 301:: 302:: 303:: 304:: 305:: Содержание
|
|
Дата добавления: 2015-07-13; Просмотров: 1202; Нарушение авторских прав?; Мы поможем в написании вашей работы!