КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Отдельные домены хроматина в политенных хромосомах могут разворачиваться и вновь упаковываться как
9-13
9.2.4. ДНК хромосом типа ламповых щеток в интерфазе состоит из серии различающихся доменов [24]
Несмотря на высокий порядок упаковки хроматина, нити его в период интерфазы слишком тонки и спутанны, чтобы можно было ясно увидеть целиком всю хромосому. Тем не менее, существуют определенные типы клеток, в которых общую структуру интерфазных хромосом различить можно. Например, спаренные в мейозе хромосомы растущих ооцитов (незрелые яйцеклетки), активно синтезируют РНК и образуют необычайно жесткие и протяженные петли хроматина, покрытые вновь транскрибируемой РНК, которая упакована в плотные комплексы РНК-
белок. В связи с тем, что ДНК покрыта такими комплексами, хромосомы (их называют хромосомами типа ламповых щеток) хорошо видны даже под световым микроскопом (рис. 9-41).

Рис. 9-41. Световая микрофотография хромосом типа ламповых щеток из ооцита земноводного. На ранних стадиях дифференцировки ооцитов каждая хромосома перед началом мейоза рсплицируется, затем гомологи спариваются с образованием четыреххроматидной протяженной структуры, изображенной на фотографии. Стадия хромосом типа ламповых щеток может длиться месяцы и даже годы. В это время в ооците создается запас мРНК и других веществ, необходимых для его последующего развития в новую особь. Обратите внимание, что каждая хромосомная ось имеет в длину приблизительно 400 мкм, тогда как длина большинства митотических хромосом составляет менее 10 мкм. (С любезного разрешения Joseph G. Gall.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 9-42. Строение хромосомы типа ламповых щеток. У многих земноводных набор хромосом типа ламповых щеток содержит в сумме около 10000 петель хроматина, хотя большая часть ДНК остается в сильно конденсированном состоянии и располагается в хромомерах. Каждая петля соответствует определенной последовательности ДНК. В каждой клетке содержится по четыре копии каждой петли, поскольку структура, представленная в верхней части рисунка, состоит из двух спаренных гомологичных хромосом, а каждая хромосома представлена двумя сестринскими хроматидами. Такие структуры, состоящие из четырех нитей, характерны для данной стадии развития ооцита (стадия диплонемы мейоза).
Схема строения хромосом типа ламповых щеток приведена на рис. 9-42. Большие петли, состоящие из деконденсированного хроматина, отходят в стороны от оси хромосомы. Опыты по гибридизации нуклеиновых кислот показали, что определенная петля всегда содержит одну и ту же последовательность ДНК, которая во время роста ооцита располагается строго определенным образом. Следовательно, эти петли соответствуют фиксированным единицам упаковки хроматина, который деконденсировался и стал транскрипционно активным. Поскольку петля среднего размера содержит приблизительно 100 000 пар оснований, каждая петля может соответствовать одной петле хроматина, описанного выше (см. разд. 9.2.1).
Многие петли постоянно транскрибируются по всей длине, другие содержат протяженные участки хроматина, который не транскрибируется вовсе.
Большая часть хроматина не входит в состав петель и остается в сильно конденсированном состоянии в хромомерах; этот хроматин, как правило, не транскрибируется. Короткие области хроматина, которые не обладают высокой степенью конденсации и активно не транскрибируются, соединяют соседние хромомеры вдоль хорошо выраженной оси хромосомы.
Хромосомы типа ламповых щеток являются необычными в том отношении, что уровень их транскрипции выше, а большинство образующихся транскриптов РНК длиннее, чем те, которые синтезируются на других хромосомах. Однако есть данные, что молекула ДНК, входящая в состав любых интерфазных хромосом, тоже подразделяется на различные области, каждая из которых отделена от своих соседей границей. По-видимому, и в данном случае хроматин в разных областях упакован по-разному (например, в виде петель, хромомеров или хроматина, входящего в состав оси между хромомерами).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
9.2.5. В политенных хромосомах также можно увидеть упорядоченные участки интерфазного хроматина [25]
Структура хроматина на уровне одиночных петель очень хорошо различима и в некоторых клетках насекомых. Многие клетки личинок мух вырастают до необычайно большого размера, претерпевая несколько циклов репликации ДНК, которые не сопровождаются клеточным делением. В результате они содержат в несколько тысяч раз больше ДНК, чем обычная клетка. Такие гигантские клетки называются полшлоидными
в том случае, если число наборов хромосом у них превышает норму. Ситуация может быть и иной: гомологичные хромосомные пары могут не отделяться друг от друга, а формировать единые огромные хромосомы (политенные хромосомы). Тот факт, что в отдельных гигантских клетках насекомых хромосомы могут переходить из политенного в по-
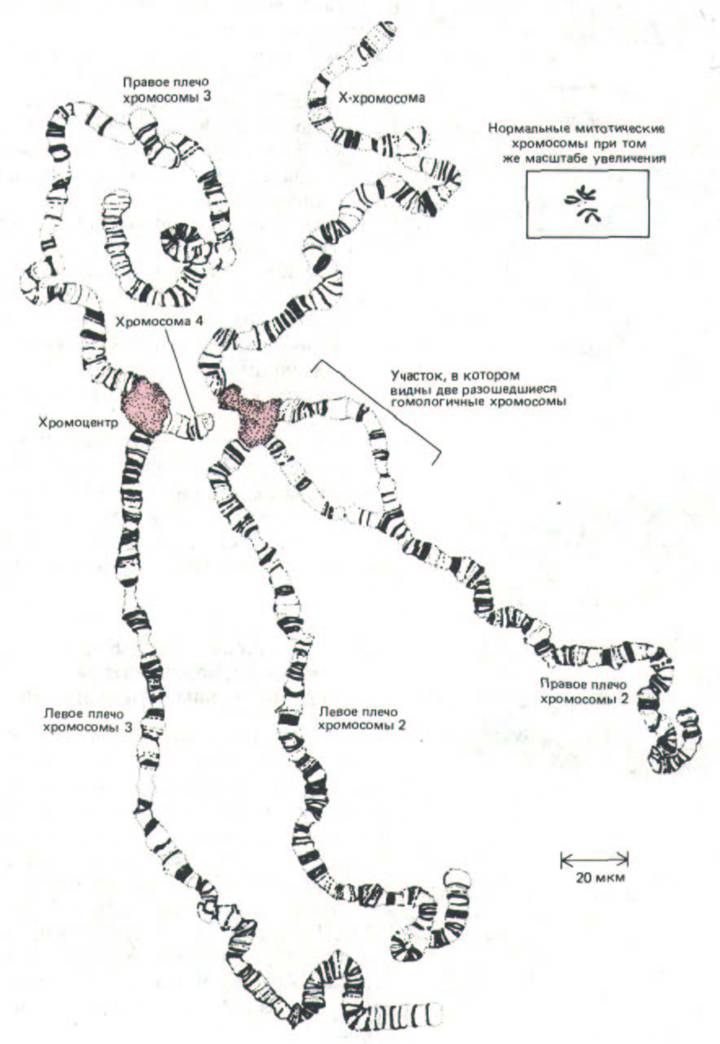
Рис. 9-43. Полный набор политенных хромосом из клетки слюнной железы дрозофилы. Эти хромосомы были расправлены и подготовлены для наблюдения путем «раздавливания» материала на предметном стекле. На рисунке представлены четыре пары хромосом. Каждая хромосома тесно спарена со своим гомологом (так, что каждая пара кажется единой структурой), чего не наблюдается в большинстве интерфазных ядер. Четыре пары хромосом связаны друг с другом своими центромерными зонами, образующими один большой «хромоцентр» (окрашенная
область). На данном препарате хромоцентр разделился на две части в процессе подготовки к микроскопированию. Необходимо отметить, что при выстраивании бок о бок многих нитей хроматина произошло значительное раскручивание каждой молекулы ДНК. (С изменениями по Т. S. Painter, J.Hered. 25: 465-476, 1934.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 9-44. Световая микрофотография участка политенной хромосомы из клетки слюнной железы дрозофилы. Видна характерная картина распределения дисков. Эти диски обнаруживаются в интерфазных хромосомах и являются отличительной чертой гигантских политенных хромосом; их не следует отождествлять с гораздо более «грубыми полосами, показанными на рис. 9-40, которые обнаруживаются в нормальных митотических хромосомах с помощью специальных красителей. (С любезного разрешения Joseph G. Gall.)

Рис. 9-45. Электронная микрофотография небольшого участка полигенной хромосомы дрозофилы. На тонком срезе видно, что разные хромосомные диски, весьма различающиеся по толщине (В), разделены междисковыми участками (I), хроматин которых гораздо менее сконденсирован. (С любезного разрешения Viekko Sorsa.)
лишюидное состояние, свидетельствует о том, что эти два хромосомных статуса тесно связаны между собой и что структура политенных хромосом в принципе аналогична структуре нормальных хромосом.
Благодаря своей большой величине, а также тому, что плотная и строго упорядоченная упаковка индивидуальных хроматшювых нитей в составе политенной хромосомы препятствует их запутыванию, эти хромосомы очень хорошо видны в световой микроскоп. Как и хромосомы типа ламповых щеток, политенные хромосомы в интерфазе активно синтезируют РНК. Явление политении наиболее глубоко было изучено на четырех хромосомах клеток слюнных желез личинок дрозофилы. В этих клетках ДНК реплицируется 10 раз подряд, дочерние хромосомы не разделяются, в результате чего образуются гигантские хромосомы, содержащие 1024 (210) тесно прилегающие друг к другу индивидуальные хроматиновые нити.
При наблюдении окрашенных политенных хромосом в световой микроскоп хорошо заметны перемежающиеся поперечные полосы:
темные (диски) и светлые (междисковые участки) (рис. 9-43 и 9-44). Каждый диск и междисковый участок состоят из 1024 идентичных последовательностей ДНК, расположенных рядом друг с другом. Около 85% ДНК в политенных хромосомах содержится в дисках и 15%-в меж дисковых участках. Хроматин каждого диска при окрашивании выглядит более темным, так как он более конденсирован, чем хроматин междисковых участков (рис. 9-45). Полагают, что диск состоит из петли, которая многократно сложена (рис. 9-46). В зависимости от размера отдельные полосы содержат от 3000 до 300000 нуклеотидных пар. Поскольку каждый диск можно идентифицировать исходя из его толщины и расположения в хромосоме, все диски могут быть пронумерованы, что дает возможность составить «карту» политенной хромосомы. Во всем геноме дрозофилы содержится примерно 5000 дисков и 5000 междисковых участков.
отдельные единицы [26]
Задолго до того, как были получены первые данные о структуре хроматина, изучение политенных хромосом позволило сформулировать гипотезу, согласно которой транскрипция генов сопровождается значительными изменениями в упаковке ДНК: отдельный диск хромосомы вздувается при активации содержащихся в нем генов и вновь конденсируется, когда гены становятся неактивными.
Для того чтобы выявить транскрибируемые области на политенной хромосоме, можно ввести в клетки радиоактивный предшественник РНК [3Н]-уридин, а затем локализовать растущий транскрипт РНК с помощью радиоавтографии (рис. 9-47). Именно таким способом было установлено, что большая часть активного хроматина находится в де-конденсированном состоянии и образует характерные хромосомные пуфы.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
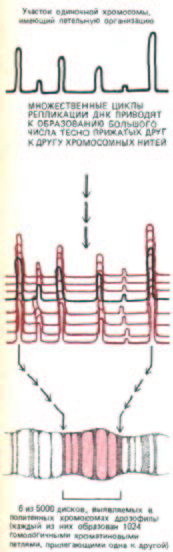
Рис. 9-46. Схема, показывающая, каким образом плотно прилегающие один к другому гомологичные петельные домены могут стать причиной появления дисков в политенных хромосомах. В составе каждого диска петли хроматиновых фибрилл находятся в очень тесном контакте друг с другом, формируя гораздо более конденсированную структуру, чем показано на этом рисунке.
Одним из основных факторов, контролирующих активность генов в политенных хромосомах дрозофилы, является гормон экдизои, встречающийся у насекомых. Уровень этого гормона в ходе развития личинки периодически поднимается и снижается, индуцируя транскрипцию разнообразных генов, которые кодируют белки, необходимые личинке для линьки и для окукливания. По мере прохождения через определенные стадии развития, возникают новые и исчезают старые пуфы, что связано с активацией и затуханием активности транскрипционных единиц и с синтезом разнообразных мРНК и белков (рис. 9-48). Изучение отдельного пуфа, размер которого относительно мал (но соответствующая ему полоса на хромосоме все-таки различима), дает основание предполагать, что каждый пуф образуется при разворачивании одного-единственного диска на хромосоме (рис. 9-49). Электронномикроскопический анализ показал, что ДНК пуфов находится в гораздо менее конденсированном состоянии, чем это свойственно хроматиновой фибрилле диаметром 30 нм (рис. 9-50). По-видимому, отдельная петля, которая, как полагают, упакована в диск на хромосоме (рис. 9-46), при транскрипции деконденсируется как самостоятельная единица.
9.2.7. Гены на политенной хромосоме расположены, вероятно, как в дисках, так и в междисковых участках [27]
Фиксированное расположение дисков и междисковых участков на политенной хромосоме дрозофилы навело цитогенетиков на мысль, что каждый диск, возможно, соответствует отдельному гену. Анализ мутаций не только подтвердил это предположение, но позволил генетикам подсчитать число жизненно важных генов у дрозофилы; их оказалось приблизительно 5000, т. е. столько же, сколько дисков на хромосомах.
Например, при попытке индуцировать мутации, картируемые на небольшом участке хромосомы (содержащем около 50 различных дисков), генетическими методами было выявлено около 50 жизненно важных генов. Хотя данный метод и не позволяет определить, где именно локализован конкретный ген, в диске или в междисковом участке, эти наблюдения дают основание для следующего вывода: обычный диск может содержать последовательности ДНК, кодирующие один-единственный жизненно важный белок.
Однако последующие эксперименты заставили усомниться в правильности гипотезы «один диск - один ген». Например, была клонирована значительная область генома дрозофилы размером 315000 нуклеотидных пар, затем отдельные ее фрагменты использовали в качестве зондов для идентификации мРНК, синтезирующихся на этом участке. Количество отдельных мРНК, как выяснилось, в три раза превышает число дисков. Весьма вероятно, что большая часть этих мРНК соответ-

Рис. 9-47. Синтез РНК на гигантской политенной хромосоме из слюнных желез насекомого Chironomus tentans. На представленном радиоавтографе хромосомы, меченной [3Н]-уридином, области синтеза РНК покрыты темными гранулами серебра в соответствии с активностью каждого участка. (По С. Felling, Chromosoma 15: 71-122, 1964.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
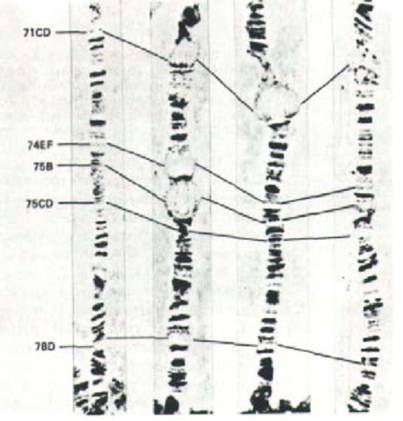
Рис. 9-48. Серия фотографий, иллюстрирующая последовательные стадии возникновения и регрессии пуфов в политенных хромосомах Drosophila melanogaster. Представлен фрагмент левого плеча хромосомы 3, содержащий пять крупных пуфов. Каждый из этих пуфов активен лишь в течение короткого периода: зафиксированная на фотографиях последовательность событий занимает 22 ч и повторяется в каждом поколении мух.
(С любезного разрешения Michael Ashburner.)

Рис. 9-49. А. Упрошенная схема процесса образования пуфа на политенной хромосоме. Б. Радиоавтограф отдельного пуфа, в котором на указанном участке синтезировалась РНК; в результате этот участок оказался меченным [3Н]-уридином. (С любезного разрешения Jose Bonner.)
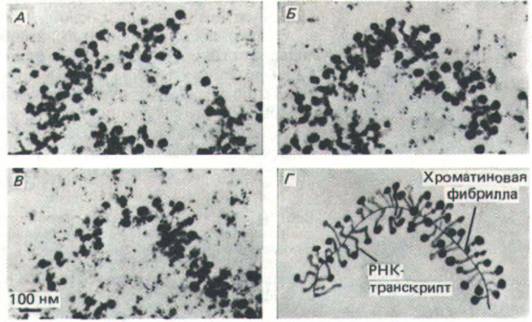
Рис. 9-50. Электронные микрофотографии серии тонких срезов крупного хромосомного пуфа, показывающие конформацию необычно длинной транскрипционной единицы. На схеме Г изображена пространственная модель исследуемого участка данной транскрипционной единицы.
Концы транскриптов РНК, похожие на клубеньки, прикреплены к одиночной хроматино-вой фибрилле. «Клубеньки» на 5'-конце большинства транскриптов образуются вследствие упаковки молекулы РНК в рибонуклеопроте-иновые частицы. (По К. Andersson, В. Bjokroth and В. Daneholt, Exp. Cell Res. 130: 313-326, 1980.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
ствуют генам, влияющим на приспособленность мух именно в природных условиях. В лабораторных условиях, в отсутствие селективного отбора, мутации по многим из этих генов могут не улавливаться. Недавно было установлено, что мРНК синтезируется как в дисках, так и в междисковых участках.
Хотя структурная организация хромосом наиболее четко выявляется при исследовании необычных интерфазных хромосом, таких как политенные или хромосомы типа ламповых щеток, вероятно, все хромосомы высших эукариот имеют подобную организацию. Изучение политенных хромосом дрозофилы показало, что гены могут располагаться и в области конденсированного хроматина дисков, и в менее конденсированных междисковых участках; кроме того установлено, что в каждом диске может содержаться несколько генов. Для чего же в таком случае каждый длинный тяж хроматина в хромосоме подразделяется на большое число различающихся областей? Хотя ответ и неизвестен, вполне вероятно, что такое строение дает возможность: I) сохранять организацию ДНК; 2} отделять гены друг от друга, и таким образом избегать биологических «помех»; 3) регулировать транскрипцию генов (например, конститутивно экспрессируемые гены «домашнего хозяйства» могут располагаться в междисковых участках, тогда как гены, специфические для определенного типа клеток, могут находиться в дисках).
|
|
Дата добавления: 2015-08-31; Просмотров: 3081; Нарушение авторских прав?; Мы поможем в написании вашей работы!