КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Связывание гистонов с ДНК приводит к образованию нуклеосом - частиц, представляющих собой единицу
|
|
|
|
9-9
9-8
9-7
9.1.14. Некоторые последовательности ДНК сильно изогнуты [12]
Спираль ДНК обладает достаточной конформационной свободой, чтобы выдерживать пружинящие изгибы и вращательные движения.
Однако в связи с тем, что эта спираль обладает и жесткостью, для того чтобы она могла изогнуться под углом 90°, в молекуле ДНК должно содержаться приблизительно 200 нуклеотидных пар. Некоторые последовательности ДНК оказываются необычайно гибкими и намного легче остальных принимают изогнутую форму.
Есть последовательности ДНК, которые склонны всегда оставаться изогнутыми. К числу последних принадлежат последовательности, в которых через каждые 10-11 нуклеотидов встречаются повторы AAAAANNNNN (где N-любой нуклеотид) (рис. 9-18). Сильный изгиб таких молекул представляет собой крайнюю форму вариаций структуры спирали. Еще не ясно, обусловлен ли такой изгиб суммарным вкладом небольших наклонов между определенными парами оснований, большим изгибом в месте соединения коротких областей или же обоими типами воздействий.
Даже если специфические последовательности, влияющие на изгиб спирали, отсутствуют, структуру ДНК в значительной мере может исказить связывание белков, принимающих участие в формировании высококонденсированных комплексов белка и ДНК.
9.1.15. Белки могут сворачивать ДНК в плотную спираль [13]
Многие белки при связывании с ДНК изгибают ее, а если подобных ДНК-белковых связей много, то нить ДНК может сформировать плотную спираль вокруг белкового комплекса с образованием нуклеопротеиновой частицы. Известно, что у бактерий такие нуклеопротеиновые частицы образуются при связывании инициаторных белков с точкой начала репликации (см. разд. 5.3.9), а также при связывании ДНК с интегразой фага лямбда для катализа сайт-специфической рекомбинации (рис. 9-19). По-видимому, в таком сложном трехмерном соединении участвуют как конкурентное, так и кооперативное взаимодействия. Аналогичные типы взаимодействий используются при регуляции каталитической активности нуклеопротеиновых частиц, как показано на примере белкового комплекса, содержащего интегразу фага лямбда (рис. 9-20). В связи с тем, что при экспрессии эукариотических генов происходит связывание кластеров белков, регулирующих активность генов,
|
|
|
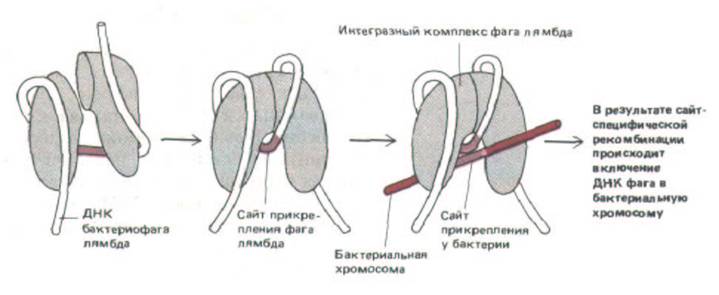
Рис. 9-19. Схема комплекса ДНК—белок, образуемого интегразой фага лямбда, ферментом, осуществляющим встраивание ДНК бактериофага в хозяйскую хромосому Е. coli. Этот комплекс катализирует сайт-специфическую рекомбинацию, разрывая и воссоединяя спирали ДНК бактериофага лямбда и бактерии по определенным участкам, называемым сайтами прикрепления (см. рис. 5-66). (С изменениями по Е. Richet, P. Abcarian, Н.А. Nash, Cell 52: 9-17, 1988.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
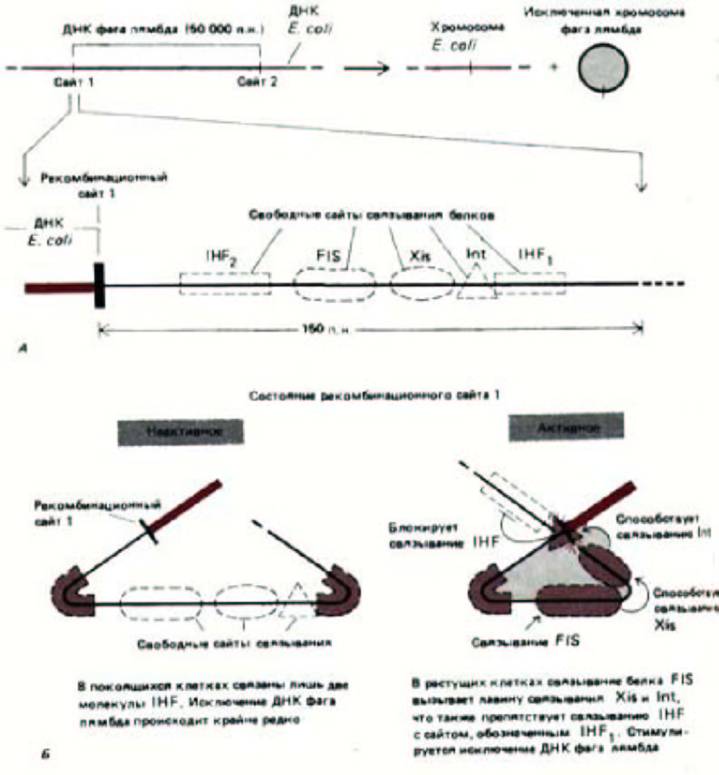
Рис. 9-20. Исключение бактериофага лямбда из хромосомы бактерии контролируется посредством кооперативного и конкурентного взаимодействия между сайт-специфическими ДНК-связывающими белками. Реакция катализируется интегразой фага лямбда и является противоположной по своему действию сайт-специфической рекомбинации, показанной на рис. 9-19. А. Общая схема реакции и некоторые участвующие в ней сайты связывания белков (указаны не все сайты). Для исключения необходимы разрыв и воссоединение двойной спирали ДНК в сайтах рекомбинации 1 и 2; при этом образуется кольцевая хромосома фага лямбда. Int-интеграза фага лямбда, Xis-эксцизионаза фага лямбда, a IHF и FIS -белки, образуемые бактериальной клеткой-хозяином. Б. Активация исключения белком FIS; указанные стадии, по-видимому, имеют место при низких концентрациях белков Int и Xis. Как показано, ряд белков при связывании сильно изгибают ДНК. Хотя белок Int катализирует реакцию сайт-специфической рекомбинации, его активность катализируется другими белками. (С любезного разрешения Arthur Landy.)
|
|
|
с определенными регуляторными последовательностями ДНК, аналогичные белковые комплексы могут принимать участие в контроле транскрипции ДНК и у эукариот (см. рис. 10-23).
Очевидно, что любые комплексы, имеющие отношение к регуляции определенных генов, должны встречаться относительно редко. В структуре хромосом эукариот присутствует главным образом другой тип нуклеопротеиновых частиц; речь идет о нуклеосоме, которая играет ведущую роль в упаковке и организации ДНК в клеточном ядре.
9.1.16. Гистоны - основные структурные белки хромосом эукариот [14]
Наиболее изученные структурные белки хромосом - это несомненно гистоны, которые имеются только в эукариотическвх клетках.
Количество их в клетках столь велико, что у эукариот принято делить белки, связывающиеся с ДНК, на два класса: гистоны и негистоновые белки
хромосом. Комплекс обоих классов белков с ядерной ДНК клеток эукариот известен под названием хроматин. Гистоны присутствуют в таких огромных количествах (около 60 млн. молекул каждого типа на клетку по сравнению с 10000 на клетку для типичного сайт-специфического белка), что их общая масса в хромосоме примерно равна содержанию ДНК.
Гистоны представляют собой относительно небольшие белки с очень
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
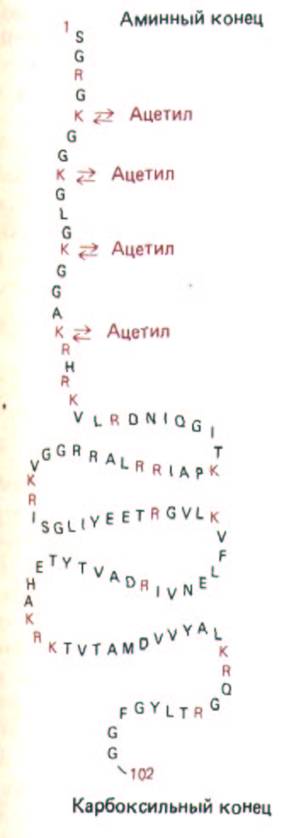
Рис. 9-21. Аминокислотная последовательность гистона Н4-одного из нуклеосомных гистонов. Аминокислоты обозначены однобуквенными сокращениями, положительно заряженные аминокислоты выделены цветом. Как и в случае других нуклеосомных гистонов, протяженная аминоконцевая последовательность молекулы обратимо модифицируется в клетке путем ацетилироваиия отдельных остатков лизина.
|
|
|
Приведенная последовательность соответствует гистону Н4 быка. У гороха этот гистон имеет почти такую же аминокислотную последовательность, за исключением того, что в нем один остаток валина замещен на изолейцин, а один остаток лизина - на аргинин.
высоким содержанием положительно заряженных аминокислот (лизина и аргинина). Суммарный положительный заряд позволяет им прочно связываться с ДНК независимо от ее нуклеотидного состава. Скорее всего гистоны постоянно находятся в комплексе с ДНК и, следовательно, играют существенную роль во всех процессах, связанных с функционированием генома.
Пять типов гистонов можно разделить на две основные группы: 1) нуклеосомные гистоны и 2) Н. гистоны. Нуклеосомные гистоны - это небольшие белки (102-135 аминокислотных остатков), отвечающие за формирование нуклеосом. К ним относятся четыре гистона: Н2А, Н2В, НЗ и Н4. НЗ и Н4 образуют внутреннюю часть нуклеосомы и, как установлено, являются наиболее консервативными из известных белков: например, аминокислотные последовательности гистонов Н4 у гороха и коровы различаются всего лишь по двум аминокислотным остаткам (рис. 9-21). Такая эволюционная стабильность предполагает, что почти каждая аминокислота, входящая в состав таких белков, играет важную роль, и изменение в любом положении может оказаться вредным для клетки.
Размер гистонов Н. больше (около 220 аминокислотных остатков), и эти белки оказались эволюционно менее стабильными, чем нуклеосомные гистоны. У дрожжей Saccharomyces cerevisiae HI, по-видимому, вообще отсутствует (см. разд. 10.3.15).
хроматина [15]
Если бы оказалось возможным растянуть нить ДНК каждой хромосомы человека, ее длина в тысячи раз превышала бы размер ядра.
Гистоиы играют важную роль в упаковке очень длинной молекулы ДНК в ядре, диаметр которого составляет всего несколько микрон. Эти белки важны и по другой причине. Известно, что ДНК может быть упакована по-разному и способ упаковки какой-либо области генома в хроматин в определенной клетке может, по-видимому, влиять на активность содержащихся в этой области генов (см. разд. 10.3.8).
|
|
|
Начало углубленному изучению структуры хроматина положило открытие в 1974 г. его основной структурной единицы-нуклеосомы.
Благодаря наличию нуклеосом частично декомпактизованныи хроматин на электронных микрофотографиях напоминает нитки бус (рис. 9-22).
«Бусину»-нуклеосому можно отделить от длинной нити ДНК путем обработки препарата хроматина ферментами, расщепляющими ДНК.
Ферменты, вызывающие деградацию как ДНК, так и РНК, называют нуклеазами, а ферменты, действующие только на ДНК,- дезоксирибонук-
леазами или ДНКазами. Нуклеаза, с помощью которой выделяют индивидуальные нуклеосомы, получена из клеток микрококков (микрококковая нуклеаза). При непродолжительной обработке этим ферментом расщепляются только те участки ДНК, которые расположены между нуклеосомами;
остальная ДНК защищена связанными с ней гистонами, вследствие чего вся молекула полимера распадается на двухцепочечные фрагменты длиной 146 пар оснований. Эти ДНК-гисто-новые комплексы на электронных микрофотографиях выглядят как частицы дисковидной формы, имеющие диаметр около 11 нм. Каждая нуклеосома содержит набор из восьми молекул гистонов - по две молекулы каждого из четырех высококонсервативных нуклеосомных гисто-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
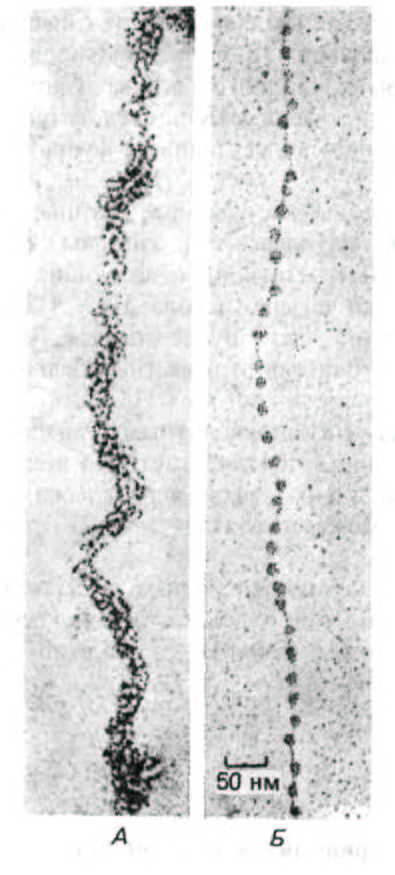
Рис. 9-22. Электронные микрофотографии хроматиновых нитей до и после обработки, приводящей к деконденсации нативной структуры и образованию «бус на нитке». А. Нативная структура, характерная для основных хроматиновых фибрилл диаметром 30 нм. Б. Деконденсированная форма хроматиновой фибриллы «бусы на нитке», показанная при том же увеличении. Схематическое изображение обеих форм хроматина приведено на рис. 9-38. Электронные микрофотографии получены с помощью модифицированной процедуры, описанной на рис. 9-71. (А-с
любезного разрешения Barbara Hamkalo, Б-с любезного разрешения Victoria Foe.)
нов: Н2А, Н2В, НЗ и Н4. Этот гистоновый октамер является по сути белковой сердцевиной нуклеосомы или ее гистоновым кором, на который накручивается фрагмент двухцепочечной ДНК (рис. 9-23).
В интактном хроматине ДНК тянется в виде непрерывной нити от нуклеосомы к нуклеосоме. Каждая нуклеосомная бусина отделена от следующей линкериой последовательностью, длина которой варьирует от 0 до 80 нуклеотидных пар. В среднем нуклеосомные частицы (нуклеосом-ный кор плюс линкерная последовательность) повторяются через каждые 200 нуклеотидов (см. рис. 9-23). Таким образом, эукариотический ген, состоящий из 10000 нуклеотидных пар, связан с 50 нуклеосомами, а в каждой клетке человека, ДНК которой насчитывает в х 109 нуклеотидных пар, содержится 3 х 107 нуклеосом.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 3122; Нарушение авторских прав?; Мы поможем в написании вашей работы!