КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Трансформация клетки в культуре позволяет выявлять гены, участвующие в социальном контроле
|
|
|
|
13-17
13.3.10. Вероятность перехода в G0 обычно увеличивается с числом клеточных делений: старение клетки [25]
У млекопитающих и птиц большинство нормальных клеток проявляет поразительную несклонность делиться неопределенно долго. Это отличает их от стабильных культивируемых клеточных линий, таких как ЗТЗ, в которых, видимо, произошли какие-то генетические изменения, делающие их «бессмертными». Например, фибробласты, взятые от человеческого плода, при выращивании в стандартной среде осуществляют только около 50 удвоений популяции; к концу этого периода пролиферация замедляется и затем останавливается, и все клетки, пробыв некоторое время в состоянии покоя, погибают. Такие же клетки, взятые от 40-летнего человека, перестают делиться примерно после 40 удвоений, а от 80-
летнего - примерно после 30 удвоений. Фибробласты от животных с более короткой продолжительностью жизни прекращают деление в культуре после меньшего числа циклов. По аналогии со старением организма в целом это было названо клеточным старением. Клеточное старение представляет собой загадочный феномен. Короткие запрограммированные серии клеточных делений, которые заканчиваются дифференцировкой, -
характерная особенность эмбрионального развития {разд. 16.3.4), однако трудно представить себе, как клетки могли бы в течение долгого времени отсчитывать свои митотические циклы и останавливаться, пройдя, скажем, 50 делений. Согласно одной из теорий, клеточное старение - это результат катастрофического накопления самовоспроизводящихся ошибок биосинтетических механизмов клетки; эти ошибки несущественны в природных условиях, где большинство животных гибнет от других причин задолго до того, как у них подвергнется старению значительное число клеток. С этой точки зрения клеточное старение просто отражает черты несовершенства в физиологии клетки, которые вполне естественны при очень слабом давлении отбора, направленного на их элиминацию. Однако в этом случае необходимо было бы объяснить, каким же образом клетки зародышевого пути, «бессмертные» клетки культивируемых линий и даже обычные соматические клетки при некоторых специальных условиях (описанных ниже) способны к бесконечной пролиферации. Другая гипотеза состоит в том, что клеточное старение-это результат механизма, который выработался для защиты от рака путем ограничения роста опухолей. Однако подобная защита представлялась бы неэффективной, так как пятидесяти циклов деления вполне достаточно
|
|
|
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
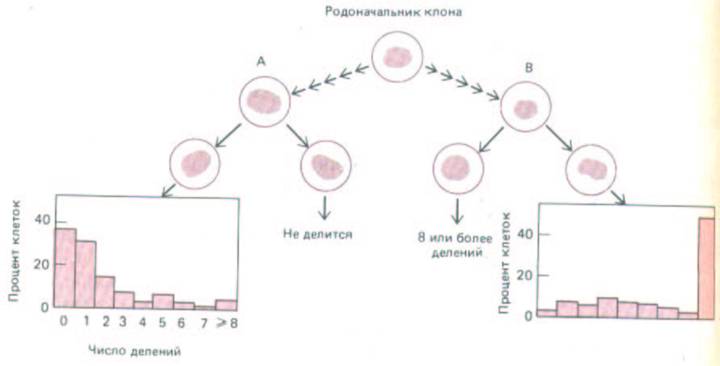
Рис. 13-30. Демонстрация различий в наследуемой способности клеток к делению. Отдельные клетки в клоне, даже будучи генетически идентичными, различаются по числу циклов деления, которые они могут осуществить. Здесь представлены разные пары сестринских клеток из одного исследованного клона, а также гистограммы, показывающие число клеток, проходящих то или иное число делений. Если одна из сестринских клеток не делится совсем, то другая обычно либо тоже не делится, либо проходит мало делений (слева); но если одна из сестринских клеток делится 8 и более раз, то другая обычно тоже претерпевает 8 и более делений (справа). Этот пример выявляет наследуемые различия между генетически идентичными клетками по числу циклов деления, которые они способны осуществить. Однако эти различные наследуемые состояния не вполне стабильны, так что иногда сестринские клетки ведут себя по-разному. Дальнейшие исследования показали, что по мере старения популяции в клетках происходят случайные переходы к пониженной способности делиться. (По данным J. R. Smith, R.G. Whitney, Science 207: 82-
|
|
|
84, 1980.)
для развития опухоли внушительных размеров. Еще одно предположение состоит в том, что старение клеток в весьма искусственных условиях клеточной культуры отражает тенденцию клеточной пролиферации в организме к постепенному замедлению с возрастом и что такое поведение клеток выработалось как способ стабилизации размеров взрослого организма.
Какова бы ни была функция клеточного старения, есть много данных о том, что на этот процесс сильно влияют факторы внеклеточной среды. Например, эпидермальные клетки из кожи ребенка стареют примерно после 50 циклов деления, если в среде отсутствует фактор роста эпидермиса, и после 150 циклов, если этот фактор имеется. «Бессмертные» клетки ЗТЗ проявляют признаки старения при недостатке факторов роста. Клетки от нормальных мышиных эмбрионов могут продолжать делиться бесконечно без малейших признаков старения, если их поместить в химическую среду определенного состава, содержащую вместо сыворотки набор очищенных факторов роста; добавление же сыворотки приводит к остановке пролиферации. Это позволяет предположить, что старение частично обусловлено какими-то компонентами сыворотки, которые тормозят пролиферацию клеток, перевешивая действие факторов роста.
Некоторые наследственные аномалии у человека, такие как синдром Вернера, приводят к преждевременному старению. Фибробласты, взятые от больных, обычно умирающих раньше 50 лет, в культуре перестают делиться после необычно малого числа митотических циклов.
Интересно то, что эти фибробласты нечувствительны к PDGF и фактору роста фибробластов, но могут энергично делиться под действием других факторов роста.
Хотя в клеточной популяции при определенных условиях старение наступает в предсказуемое время, на уровне отдельной клетки оно не является жестко запрограммированным. В клоне по видимости идентичных нормальных фибробластов, растущих в стандартных условиях, одни клетки делятся многократно, тогда как другие - всего несколько раз. Индивидуальные клетки, по-видимому, перестают делиться в результате случайного перехода в иное состояние, вероятность которого возрастает в каждом последующем поколении клеток, пока не наступит момент, когда в популяции совсем не останется делящихся клеток (рис. 13-30).
|
|
|
Исследования на клеточных клонах показывают, что клетки, которые во всех других отношениях идентичны, различаются по способности к делению. Видимо, переход стареющей клетки в непролиферирующее состояние - это просто конечный результат ряда случайно распреде-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
ленных во времени этапов ухудшения экспрессии генов, регулирующих готовность клеток проходить точку рестрикции. Молекулярная природа таких событий начинает проясняться благодаря исследованию раковых клеток, которые, помимо прочего, «бессмертны» и не подвержены старению.
Заключение
Клеточное деление у многоклеточных животных зависит от сложных «социальных» регуляторных механизмов, и пролиферация
различных типов клеток контролируется различными сочетаниями белковых факторов роста. Они действуют в очень малых концентрациях, и
многие из них служат локальными химическими медиаторами, позволяющими регулировать плотность клеточной популяции. Кроме того, большинство нормальных клеток неспособно делиться без прикрепления к внеклеточному матриксу. При недостатке факторов роста или при
невозможности прикрепиться к матриксу клетки останавливаются после митоза, переходя в особое состояние покоя —G0 из которого после
добавления факторов роста они могут выйти лишь через несколько часов. Когда клетка вышла из состояния G0 и прошла точку рестрикции в G1, она быстро проходит фазы S, G2 и М независимо от прикрепления или факторов роста. В пролиферирующей клеточной популяции переход через
точку рестрикции представляет собой событие типа «всё или ничего», которое, подобно радиоактивному распаду, характеризуется
|
|
|
определенной вероятностью осуществления. В дополнение к непосредственному контролю клеточной пролиферации существуют еще
долговременные механизмы, приводящие к старению и прекращению деления нормальных соматических клеток млекопитающих в культуре после
ограниченного числа циклов деления.
13.4. Гены «социального контроля» клеточного деления [26]
Как мы видели на примере дрожжей, генетика располагает большими возможностями для выяснения молекулярных основ регуляции клеточного деления, если имеются способы отбора мутаций соответствующих генов. У многоклеточных животных мутации генов, непосредственно участвующих в «социальном контроле» клеточного деления (мы будем называть их генами социального контроля), выделять нетрудно. Клетка, содержащая одну или несколько таких мутаций, будет продолжать делиться, игнорируя нужды организма как целого, и ее потомство образует макроскопически видимую опухоль.
Раковые опухоли - по определению злокачественные, т. е. их клетки не только делятся неконтролируемым образом, но и проникают в другие ткани, где появляются многочисленные вторичные опухоли, или метастазы. Для возникновения рака необходимо, чтобы в клетке сначала произошел ряд мутаций, освобождающих ее от воздействия различных регуляторов клеточного деления, а затем накопились дальнейшие изменения, делающие ее способной к инвазии и метастазированию. Эти аспекты рака будут обсуждаться в гл. 21. Здесь же мы не будем пытаться объяснить природу рака, а посмотрим, что можно узнать при изучении раковых клеток относительно тех генов, которые в норме контролируют клеточное деление.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 557; Нарушение авторских прав?; Мы поможем в написании вашей работы!