КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Некоторые протоонкогены кодируют внутриклеточные медиаторы, участвующие в стимуляции деления
13-22
13-16
13-21
13-20
13.4.2. Опухолеродные вирусы служат источником легко клонируемых онкогенов [28]
Опухолевый вирус разрушает нормальный контроль клеточного деления, необратимо изменяя генетическую конституцию клетки-
хозяина; в результате этого клетка начинает вырабатывать белок, не подверженный воздействию нормальных регуляторных механизмов. Поэтому такие вирусы дают возможность выявлять механизмы, ответственные в норме за контроль клеточного деления. До сих пор наиболее важные результаты были получены при изучении РНК-содержащих опухолевых вирусов, называемых также ретровирусами. После заражения клетки ретровирусом на его РНК путем обратной транскрипции синтезируется ДНК, которая затем включается в геном клетки-хозяина. Жизненный цикл ретровируса представлен на рис. 5-75.
Когда ретровирус трансформирует нормальную клетку в опухолевую, аномальное поведение часто бывает обусловлено геном, который (
привнесен вирусом, но для выживания и репродукции самого вируса фактически не нужен. Впервые это выяснилось, когда были открыты ' мутанты вируса саркомы Рауса, способные нормально размножаться, но не трансформирующие клетку. Оказалось, что некоторые из этих нетрансформирующих мутантов не имеют гена (или части гена), кодирующего белок с мол. массой 60000. В результате других мутаций этого гена трансформирующее действие вируса может становиться термочувствительным: зараженные клетки проявляют трансформированный фенотип при 34°С, но после повышения температуры до 39°С они быстро (через несколько часов) возвращаются к нормальному фенотипу (рис. 13-32). По-
видимому, этот специфический ген в онкогенном вирусе ответствен за клеточную трансформацию (и этим привлекает наше внимание), но является ненужным балластом с точки зрения репродукции самого вируса.
Трансформирующий ген вируса саркомы Рауса, выявленный в этих
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
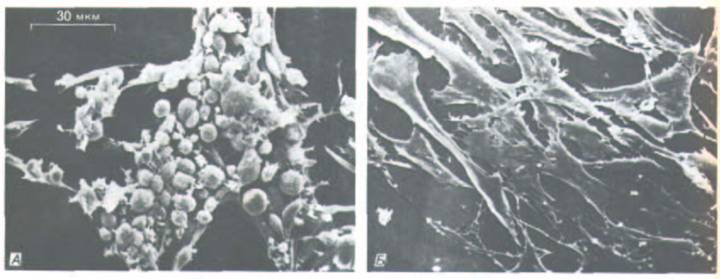
Рис. 13-32. Клетки, инфицированные вирусом саркомы Рауса, несущего термочувствительную мутацию гена, ответственного за трансформацию (онкогена v-src) (микрофотографии, полученные с помощью сканирующего электронного микроскопа). А. Клетки трансформированы и приобрели аномальную округлую форму при низкой температуре (34 °С), при которой продукт онкогена активен. Б. Те же клетки, прочно прикрепленные к культуральной чашке и восстановившие свою нормальную уплощенную форму, после того как продукт онкогена инактивирован повышением температуры (до 39 °С). (С любезного разрешения G. Steven Martin.)
экспериментах, был назван геном v-src. Он был отнесен к онкогенам (от греч. onkos-масса, опухоль), так как при введении в нормальную клетку он превращает ее в опухолевую. Каково происхождение этого гена и какова его нормальная функция? Когда радиоактивная ДНК-копия вирусного гена src была использована как зонд для поиска родственных последовательностей методом ДНК-ДНК-гибридизации (разд. 4.6.7), оказалось, что геномы нормальных клеток позвоночных содержат очень сходную, но не идентичную последовательность. Этот гомолог вирусного гена src в нормальной клетке обозначают c-src и относят к протоонкогенам. По-видимому, вирусный онкоген был когда-то захвачен из генома клетки-хозяина и подвергся мутации. Можно предполагать, что протоонкоген - это один из нормальных генов социального конгроля, а ретровирус, по существу, клонировал его для нас. В настоящее время таким же способом выявлено и проанализировано много других онкогенов, и в каждом случае это привело к открытию соответствующего протоонкогена.
13.4.3. Опухоли, возникающие разными способами, содержат мутации одних и тех же протоонкогенов [26, 29]
Опухоли часто возникают не от вирусной инфекции, а в результате мутаций, происходящих случайно или же под действием химических канцерогенов или облучения. Из таких опухолевых клеток можно получить очищенную ДНК и проверить ее на наличие онкогенов путем введения в нетрансформированные клетки в культуре in vitro. Для такого теста часто используют клетки ЗТЗ, поскольку они неопределенно долго делятся в культуре и содержат мутации, благодаря которым их легко трансформировать добавлением всего лишь одного онкогена. Онкоген, ответственный за такую трансформацию, может быть выделен, клонирован и секвенирован с помощью метода рекомбинантной ДНК (рис. 13-33). Примечательно, что в большинстве случаев, когда это было сделано, онкоген оказывался мутантной формой одного из тех самых протоонкогенов, которые были выделены с использованием ретровируса, хотя при этом было открыто и несколько новых онкогенов.
При помощи сходных методов было установлено, что трансформация может также происходить в результате перепроизводства некоторых нормальных генных продуктов. Часто опухоли содержат неизмененный протоонкоген, избыточная экспрессия которого обуслов-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
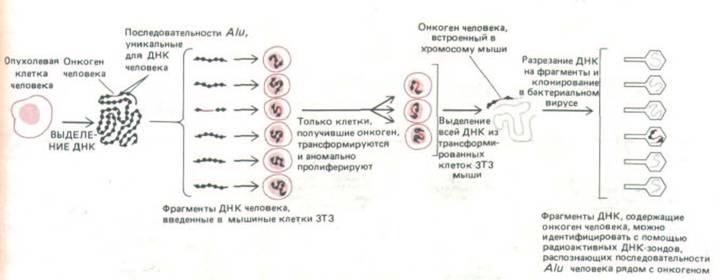
Рис. 13-33. Метод, позволяющий идентифицировать и клонировать онкогены человека. Онкогены, содержащиеся в образце ДНК, взятом из опухоли человека, выявляются по их способности трансформировать мышиные клетки ЗТЗ. Трансформированные клетки ЗТЗ безудержно делятся и распознаются по колониям, которые они образуют в культуральной чашке. Повторяющиеся последовательности семейства Alu (разд.
10.5.11) разбросаны по всему геному человека и служат удобным маркером, который можно использовать для идентификации ДНК человека в клетке другого организма. Используемые в качестве ДНК-зонда последовательности Alu позволяют клонировать онкоген человека, трансформирующий клетки ЗТЗ. При повторном тестировании этим же методом клонированная ДНК, содержащая онкоген, будет очень эффективно трансформировать клетки ЗТЗ.
лена либо тем, что он представлен слишком большим числом копий, либо тем, что хромосомная перестройка поставила его под контроль несоответствующего промотора. Эти вопросы будут обсуждаться в гл. 21.
До сих пор идентифицировано более 50 протоонкогенов (разд. 21.2.3, 21.2.4). Они, видимо, составляют значительную часть протоонкогенов нормальной клетки. Однако многие гены социального контроля, вероятно, еще не обнаружены. Фибробластоподобные клетки ЗТЗ, обычно используемые в тестах на трансформацию, могут не поддаваться действию того онкогена, который трансформировал какие-то другие виды дифференцированных клеток. Кроме того, тест на клеточную трансформацию позволяет выделять только доминантные мутации генов социального контроля, т. е. такие мутации, которые нарушают регуляцию клеточного деления даже при наличии в клетке копий нормального аллеля. Возможно, что в раковых клетках чаще встречаются рецессивные мутации генов социального контроля, обусловленные утратой генной функции, но их нельзя выявить в таком тесте. Поэтому гены, продукты которых в норме помогают стимулировать деление клеток, легче идентифицировать с помощью современных методов, чем те, продукты которых в норме тормозят деление. Тем не менее уже есть данные о том, что гены с тормозящим действием существуют и что их рецессивные мутации нередко бывают причиной клеточной трансформации и рака. Например, после слияния трансформированных клеток с нетрансформированными полученные гибридные клетки очень часто ведут себя как нормальные; по-видимому, контроль клеточного деления у них восстанавливается благодаря появлению белка, которого не было у трансформированной клетки. Поэтому, помня о том, что многие важные гены социального контроля еще не открыты, мы теперь рассмотрим функции уже известных генов.
13.4.4. Некоторые протоонкогены кодируют факторы роста или рецепторы факторов роста [26, 30]
Если онкоген клонирован и секвенирован, то указания на вероятную функцию соответствующего гена социального контроля (протоонкогена) часто можно найти, сравнивая последовательности нуклеотидов данного онкогена и уже известных генов. Именно таким путем было открыто,
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
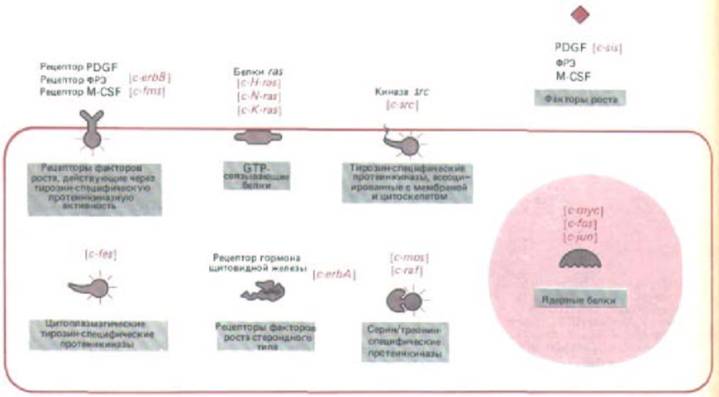
Рис. 13-34. Активность и локализация в клетке основных классов известных протоонкогенов. Названия некоторых типичных протоонкогенов каждого класса выделены цветом. См. также рис. 21-27.
что один из протоонкогенов, c-sis, кодирует функционально активную субъединицу фактора роста PDGF. Клетка, содержащая соответствующий онкоген v-sis, непрерывно и без надобности вырабатывает эту субъединицу, и она, связываясь с клеточным рецептором для PDGF, все время побуждает клетку к пролиферации. По крайней мере три других протоонкогена - c-erbB, c-fms и с-еrbА - тоже кодируют рецепторы для факторов роста или гормонов: с-erbB кодирует рецептор для фактора роста эпидермиса (разд. 12.3.13), c-fins- рецептор фактора, стимулирующего рост колоний макрофагов (M-CSF) (этот фактор способствует пролиферации предшественников макрофагов, разд. 17.5.8), a c-erbA- рецептор гормона щитовидной железы (разд. 12.2). Превратившись в результате мутаций в онкогены, эти гены будут кодировать измененные рецепторы, которые ведут себя так, как будто присоединили лиганд (даже если его нет), и поэтому стимулируют клетку, когда это не нужно (разд. 12.3.14).
Хотя функции большинства других протоонкогенов еще в точности не известны, можно предполагать, что большинство из них кодирует белки внутриклеточной сигнальной сети, дающей возможность факторам роста стимулировать клеточную пролиферацию. Теперь мы должны рассмотреть, насколько широкий диапазон функций могут осуществлять уже известные группы протоонкогенов, представленные на рис. 13-34.
|
|
Дата добавления: 2015-08-31; Просмотров: 554; Нарушение авторских прав?; Мы поможем в написании вашей работы!