КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Історія клонування
|
|
|
|
Всі клітини організму тварин несуть однакову генетичну інформацію. Однак у процесі морфогенезу соматичні клітини диференціюються, внаслідок чого частина генома репресується. Чим вищий рівень спеціалізації клітин, тим менша їх тотипотентність. Ця закономірність була встановлена в експериментах з пересадження ядер.
Уперше трансплантацію ядер соматичних клітин зародків до енуклеїованих клітин жаби здійснили американські дослідники Р. Бриггс і Т. Кінг у 1952 році (рис. 59). Учені, користуючись мікропіпеткою, видаляли ядра з яйцеклітин шпорцевої жаби, а замість них пересаджували ядра клітин ембріонів, що перебувають на різних стадіях розвитку. Проведені дослідження показали, що ядра ранніх ембріонів у стадії пізньої бластули й навіть ранньої гаструли володіють тотипотентністю й забезпечують нормальний розвиток ембріонів. Якщо брати ядра із клітин зародка на ранній стадії його розвитку - бластули, то майже у 80% випадків зародок нормально розвивається далі і перетворюється на справжнього пуголовка. Якщо ж розвиток зародка, донора ядра, переходить на наступну стадію -гаструлу, то лише менш ніж у 20% випадків оперовані яйцеклітини розвивалися нормально. При пересадженні ядер з більш диференційованих клітин (мезодерми й середньої кишки) пізньої гаструли в ембріонів спостерігалося недорозвинення і навіть відсутність нервової системи. Після пересадження ядра із клітин більш пізнього розвитку яйцеклітини взагалі не розвивалися.
Більш ґрунтовні дослідження, що охоплюють не тільки амфібій, а й риб, а також дрозофіл, у 1962 р. були розпочато англійськимбіологом Дж. Гордоном. Він першим у дослідах зпівденноафриканськими жабами (XenopusLaevis) як донора ядервикористав не зародкові клітини, а клітини епітелію кишечникаплаваючого пуголовка, що вже цілком спеціалізувалися. Ядраяйцеклітин реципієнтів він не видаляв хірургічним шляхом, аруйнувавультрафіолетовимипроменями.Здебільшогореконструйовані яйцеклітини не розвивалися, але приблизно десята частина з них утворювала ембріони. 6,5% із цих ембріонів досягали стадії бластули, 2,5% − стадії пуголовка і тільки 1% розвився в статевозрілих особин. Однак, поява декількох дорослих особин у таких умовах могла бути пов'язана з тим, що серед клітин епітеліюкишечника пуголовка, що розвивається, досить тривалий час присутні первинні статеві клітини, ядра яких могли бути використані для пересадження. У наступних роботах як сам автор, так і багато інших дослідників не змогли підтвердити дані цих перших дослідів.
|
|
|
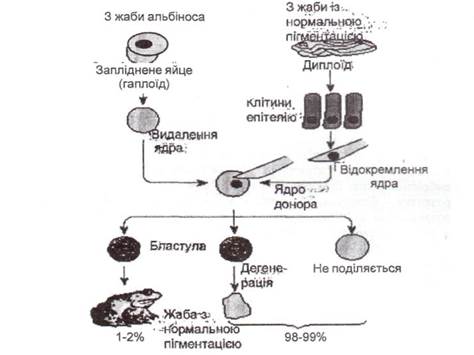
Рис. 59. Схема отримання клонованої жаби
У подальшому Дж. Гордон разом з Пещення (1970) почали культивувати invitro клітини нирок, легень та шкіри дорослих тварин і використовувати вже ці клітини як донори ядер. Майже 25% первинно реконструйованих яйцеклітин розвивалися до стадії бластули. При серійних пересадженнях вони розвивалися до стадії плаваючого пуголовка. У такий спосіб було показано, що клітини трьох різних тканин дорослого хребетного (X. Lаvуіs) містять ядра, які можуть забезпечити розвиток принаймні до стадії пуголовка.
У свою чергу М. Дж. Берардино і Н.Хофнер(1983) використовували для трансплантації ядра клітин крові, що не поділяються та цілком диференційовані − еритроцити жаби Rапаріріеs. Після серійного пересадження таких ядер 10% реконструйованих яйцеклітин досягали стадії плаваючого пуголовка. Ці експерименти показали, що деякі ядра соматичних клітин здатні зберігати тотипотентність.
У 1985 р. була описана технологія клонування кісткових риб, що розроблена радянськими вченими Л.А. Слєпцовою, Н.В. Дабагян і К.Г. Газарян. Зародки на стадії бластули відокремлювали від жовтка. Ядра клітин зародків вприскували в цитоплазму незапліднених ікринок, які починали дробитися й розвивалися в личинки. Ці експерименти показали, що втрата ядром тотипотентності в процесі онтогенезу пов'язана не з втратою генів, а їхньою репресією. За культивування соматичних клітин іп vitro частота тотипотентності ядер збільшується.
|
|
|
Пересадження ядер у ссавців почалися пізніше, у 80-х роках. Це було пов'язано з технічними труднощами, тому що зигота ссавців має невеликі розміри. Наприклад, діаметр зиготи миші приблизно 60 мкм, а діаметр заплідненої яйцеклітини жаби − 1200 мкм, тобто у 20 разів більше. Зигота корові трохи більша за зиготу миші, діаметр її становить 160 мкм, але пронуклеуси приховані яєчним жовтком, тому перед мікроманіпуляціями необхідна спеціальна обробка зигот.
Незважаючи на названі труднощі, перші повідомлення про одержання клонів мишей, ідентичних донору, з'явилися вже у 1981 році. Як донори були використані ембріональні клітини однієї з ліній мишей, узяті на стадії бластоцисти. Вірогідність отриманих даних спочатку була поставлена під сумнів тому, що відтворити результати проведених експериментів у інших лабораторіях не вдавалося, однак через два роки Дж. Мак Ґрат і Д. Солтер також досягай успіху. Американські дослідники С. Стік і Дж. Робл, використовуючи методику Мак Грата й Д. Солтера, у 1988 р. отримали шість живих кролів, пересадивши ядра 8-клітинних ембріонів однієї породи в позбавлені ядра яйцеклітини кролів іншої породи. Фенотип народжених повністю відповідав фенотипу донора. У цих експериментах тільки 6 з 164 реконструйованих яйцеклітин (3,7%) розвилися в нормальних тварин.
Перші успішні експерименти з клонування сільськогосподарських тварин були проведені С. Уілладсеном (S. Wіlladіsеп) у 1986 р. Він зливав без'ядерні яйцеклітини із бластомерами,виділеними з 8- і 16-клітинного ембріона вівці.
Дж. Робл і його співробітники у 1987 р. провели роботи з пересадження ядер великої рогатої худоби. Вони пересаджували до зиготкаріопласти −чоловічий і жіночий пронуклеуси разом з цитоплазмою, що їх оточує, а також ядра 2-, 4- або 8-клітинних ембріонів корови. Реконструйовані зародки в цій роботі розвивалися тільки в тих випадках, коли до зигот пересаджували пронуклеуси: 17% таких зародків досягли стадії морули або бластоцисти. Два зародки були пересаджені іншому реципієнтові − у матку корови, і розвиток їх завершився народженням живих телят. Якщо як донорів використовували ядра 2-, 4- або 8-клітинних зародків, то реконструйовані яйцеклітини не розвивалися навіть до стадії морули. Пізніше були й більш успішні роботи. Зокрема С. Уілладсен (1989) повідомив, що йому вдалося отримати чотирьох генетично ідентичних бичків голштинської породи пересадженням до реципієнтних яйцеклітин ядер бластомерів одного 32-клітинного зародка.
|
|
|
К. Бондіолі й співавтори (1990), використовуючи як донорів ядер 16-64-клітинні зародки корів, трансплантували 463 реконструйованих зародки в матку синхронізованих реципієнтів, було отримано 92 живих теляти. Сім з них були генетично ідентичні, являючи собою клон, отриманий у результаті пересадження ядер клітин одного донорського ембріона.
Експериментів з клонування свиней небагато. Успішні дослідження провели Р. Пратер зі співробітниками в 1989 р. Незначна кількість даних пов'язана з певними труднощами роботи із цим об'єктом.
У 1993... 1995 роках група дослідників під керівництвом Я. Уілмута (LапWіlтиt) з Рослинского інституту отримала клон овець - 5 ідентичних тварин, донорами ядер яких була культура ембріональних клітин.
Я. Уілмут зі співавторами опублікував на початку 1997 року повідомлення, що в результаті використання донорського ядра клітини молочної залози вівці було отримано клоновану тварину-вівцю за кличкою Доллі.
Аналогічні експерименти здійснювали пізніше Т. Домінко (ТаnjaDоmіпко) і співробітники лабораторії Вісконсинського університету, які забезпечили клонування ембріонів із клітин шкіри вух дорослої рогатої худоби. Ембріони, генетично ідентичні корові,що пожертвувала клітини вуха, були пересаджені в матки корів-реципієнтів. Спостерігалася поступова загибель ембріонів, тому життєздатних телят не отримали. Причини поки що не встановлено.
У серпні 1997 року з'явилося повідомлення про те, що Алан Троунсон (Австралія) розробив технологію, яка дозволяє сформувати ембріон з 16, 32 або 64 клітин, а потім кожна з них може використовуватися для формування 16, 32 або 64 ідентичних ембріонів. Колектив дослідників на чолі з Аланом Троунсоном створив 470 генетично ідентичних ембріонів рогатої худоби від єдиної бластоцисти. Така технологія забезпечує безмежне джерело генетичного матеріалу для клонування.
|
|
|
|
|
Дата добавления: 2017-02-01; Просмотров: 184; Нарушение авторских прав?; Мы поможем в написании вашей работы!